Ella Hu, Siri Jayaprakash, Sanjith Kalpat, Tomas Kruecker-Green, and Keerthi Shome, Andover High School, Andover, Massachusetts, United States
Reviewed on 3 May 2025; Accepted on 9 June 2025; Published on 27 October 2025
With help from the 2025 BioTreks Production Team.
Every year, approximately 400 million metric tons of plastic are produced globally, with polyethylene terephthalate (PET) contributing 14% to that amount. Only 5% of the PET is recycled, 15% is incinerated, and 75% is landfilled. Recently, enzymes such as PETase, from Ideonella sakaiensis, and leaf compost cutinase (LCC), found in compost piles, have shown potential in addressing this long-standing issue. However, the temperature of piles in recycling facilities is often 55-60°C, so a thermostable enzyme like LCC is needed. Two different forms of LCC were observed, one with the original sequence and another engineered to be more thermostable. The engineered LCC ultimately worked better compared to the original LCC. The efficiency of PETase and LCC against PET film (0.25 mm thickness) and PET powder (300 μm) was evaluated. Both enzymes were expressed in a cell-free system (BioBits®) by adding DNA at 100 ng/µL, and using green fluorescent protein (GFP) as a reporter. The level of expression was inferred using ImageJ to quantify fluorescence. The enzymes were then applied to both the PET film and powder and incubated for five days. Imaging with scanning electron microscope (SEM) indicated that PET films with engineered LCC showed decomposition, and PET powder particles with engineered LCC had smoother and rounder edges when compared with negative controls.
Keywords: PET, polyethylene terephthalate, PETase, LCC, leaf compost cutinase, degradation of plastic
Authors are listed in alphabetical order. Lindsey L'Ecuyer and Nathan Crook mentored the group. Please direct all correspondence to nccrook@ncsu.edu.

Background
Over 430 million tons of plastic are produced globally each year, yet only 9% of the plastic waste is being recycled (Organisation for Economic Co-operation and Development [OECD], 2022). Among the most widely used plastics is polyethylene terephthalate (PET), a thermoplastic polymer commonly found in plastic bottles. PET alone is responsible for roughly 10.2% of global plastic production. Most discarded PET ends up incinerated, in landfills, or in the ocean, harming marine life.
PETase (from Ideonella sakaiensis) breaks down PET by hydrolyzing the ester bonds in the polymer, converting it into intermediate products like Bis(2-Hydroxyethyl) terephthalate (BHET) and mono(2-hydroxyethyl) terephthalate (MHET). PETase is a member of the α/β-hydrolase fold family and contains a catalytic triad (Ser-His-Asp) in its active site, which facilitates this hydrolysis reaction. The process occurs in two steps; first, the enzyme forms an acyl-enzyme intermediate with the PET substrate; second, this intermediate is hydrolyzed, releasing MHET and other products.
Leaf-branch compost cutinase (LCC) is a thermostable cutinase discovered in hot compost. Unlike PETase, LCC can depolymerize PET almost completely in one step. In vitro, LCC treatment of PET film yielded essentially only TPA (major) and EG, with only trace MHET and no detectable BHET (Suliman et al., 2012). This indicates that LCC hydrolyzes PET directly to its monomeric components (TPA and EG) without accumulating BHET intermediaries, thus “bypassing” the typical MHET bottleneck. Instead, LCC is able to cleave PET hydrolyzed products quickly so that the final monomers accumulate. Crucially, LCC and its engineered variants are highly thermostable. For example, optimized LCC variants have the most activity around 70-80°C and retain roughly half their activity even after hours at 90°C.
Materials and methods
Designing and obtaining PETase and LCC (original) and LCC (modified) vectors
Three plasmid vectors were designed in Benchling and ordered from Twist Biosciences, each containing the gene sequence for PETase, LCC (original), or LCC (modified), linked to green fluorescent protein (GFP) and integrated into the pJL1 backbone (Figures 1-3). LCC (modified) was known to be more thermostable, based on work in the lab of mentor Dr. Nathan Crook. All sequences were codon optimized for expression in Escherichia coli using a ribosome binding site (RBS)calculator and parameters defined by Twist. These vectors were designed for expression in a cell-free protein synthesis system called BioBits® (miniPCR, 2019; Stark et al., 2018). The constructs contain a T7 promoter terminator chosen to function specifically with the BioBits® cell-free expression system, which requires strong genetic parts for high affinity to the cell-free ingredients. They also contain strong RBS and a His-tag on the GFP. The vectors were ordered in TE buffer. Twist could not normalize the concentrations, so the resulting DNA arrived at different concentrations (PETase 471.42 ng/µL, LCC 371 ng/µL).
| Figure 1. pJL1 + PETase + GFP Plasmid. |
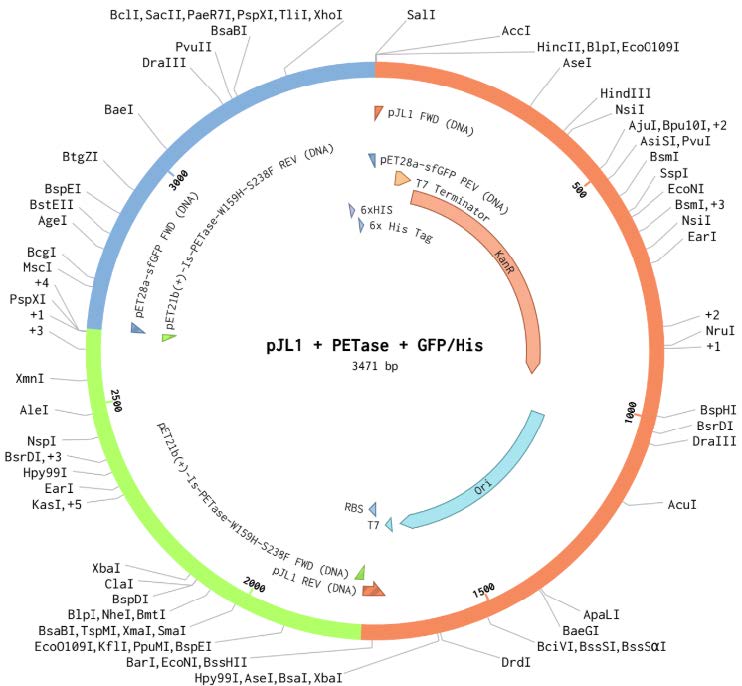
|
|---|
| Figure 2. pJL1 + LCC (original) + GFP Plasmid. |
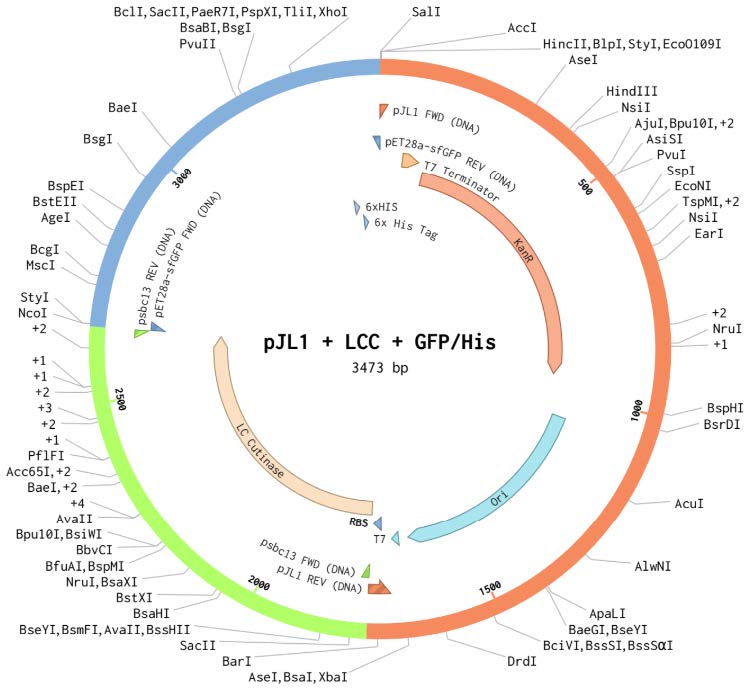
|
|---|
| Figure 3. pJL1 + LCC (modified) + GFP Plasmid. |
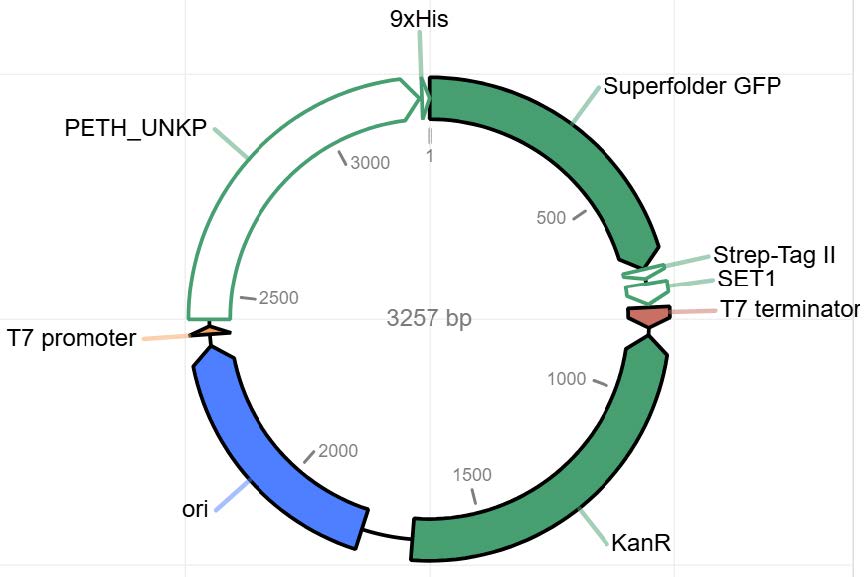
|
|---|
Evaluating enzymatic activity of PETase and LCC through GFP expression in BioBits®
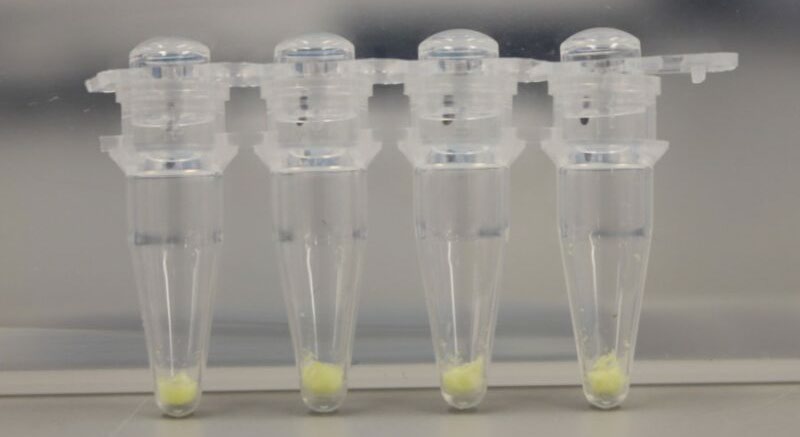
BioBits®, a freeze-dried cell-free protein expression system (Figure 4), was obtained from miniPCR.
| Figure 4. BioBits® (miniPCR, 2019). |
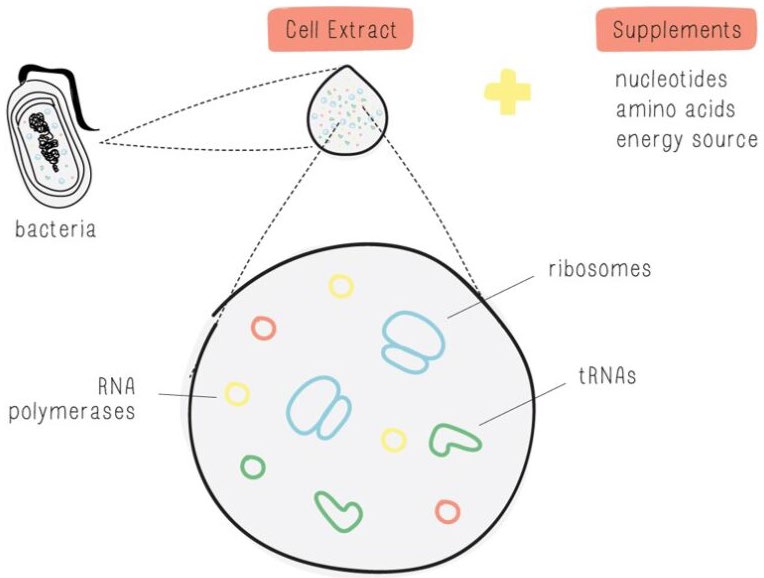
|
|
|---|
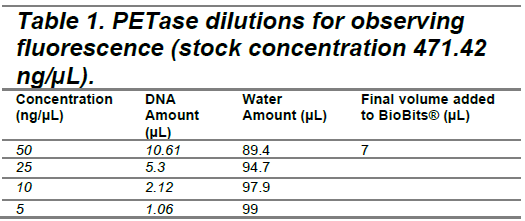
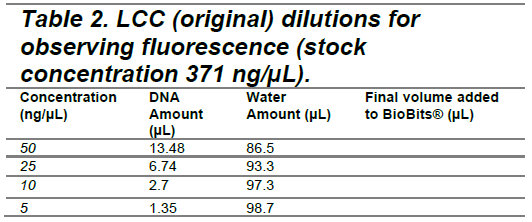
The PETase and LCC (original) plasmids were added to BioBits® reactions at concentrations of 50 ng/µL, 25 ng/µL, 10 ng/µL, and 5 ng/µL (Tables 1 and 2) to determine the optimal DNA concentration for fluorescence visualization. Following DNA addition, fluorescence images were captured at hourly intervals over a 9-hour period to monitor GFP expression as an indicator of protein synthesis. A plasmid encoding red fluorescent protein (provided in the BioBits® kit) served as a positive control, while nuclease-free water was used as a negative control (Tables 3 and 4). Following the experiment, differences in GFP expression were assessed between each experimental vector and the negative control to evaluate relative protein production. The protein production shown by GFP fluorescence should be qualitatively visible; however, ImageJ software can also be used to quantify fluorescence intensity and confirm observations.
|
|---|
|
|---|

|
|---|

|
|---|
Applying PETase and LCC to PET films and powder substrates to determine the ability of the enzymes to break down plastic
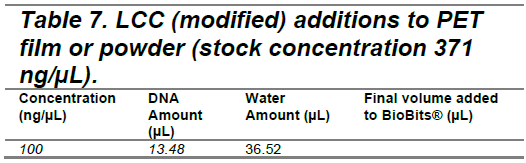
Following our initial experiment assessing the enzymatic activity of PETase and LCC (original), follow-up experiments were conducted using a modified version of LCC engineered for enhanced thermostability. DNA from the three vectors was added to separate BioBits® reactions at 100 ng/µL and allowed to incubate to allow time for expression of the enzymes (Tables 5 -7). The enzymes were applied to the biaxially oriented PET films at 0.5 mm in thickness and semi-crystalline (crystallinity>40%) powder substrates with a maximum size of 300 µm. The film and powder were obtained from Dr. Nathan Crook and Dr. Tianyu Li’s lab at North Carolina State University (NCSU). The enzymes with film or powder were then incubated for five days. Samples containing PETase were incubated at room temperature (22°C), while those containing the original and modified LCC were incubated at 72°C to match their optimal operating temperatures.
Afterwards, PET films were rinsed three times each with 20µL of distilled water, and PET powders were rinsed three times with 40µL of distilled water in order to remove any DNA or residual BioBits®. PET powders were rehydrated by warming in 40µL of distilled water for ten minutes and briefly spinning in a centrifuge.
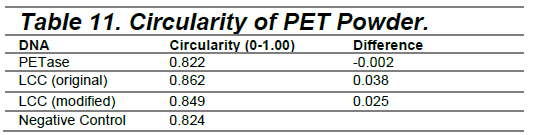
To evaluate the degradation, a scanning electron microscope (SEM) was used. Samples were prepared for SEM imaging by drying overnight in a desiccator and followed by a gold coating using a sputter coater (Denton Vacuum Desk IV). The sputtering process lasted for two minutes, resulting in a 10 μm coating. The PET films and powder substrates were imaged at 50X, 250X, and 1000X magnification. Subsequently, ImageJ was used to analyze the circularity of the PET powder particles in order to quantitatively assess the extent of enzymatic degradation

|
|---|

|
|---|
|
|---|

|
|---|
Assessing enzymatic degradation in PET powder through ImageJ
Using ImageJ, fluorescent particles were isolated from the background by selecting “Process,” followed by “Find Edges.” Threshold settings were then adjusted to highlight the particles of interest and maintain a consistent threshold value across all analyzed scans to ensure comparability. After thresholding, “Analyze Particles” function was used to extract data, including the average circularity and size of the particles in each scan.
Results
PETase vectors show strong GFP expression, LCC vectors do not
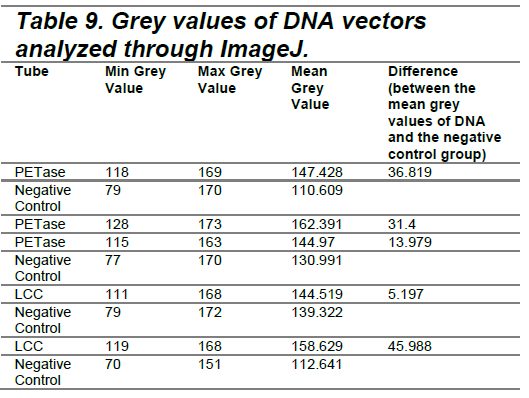
The data indicated that the vectors containing PETase and LCC demonstrated measurable protein production, shown by GFP fluorescence after 8 hours. Observations were quantified using ImageJ software by measuring the minimum and maximum grey values (Table 9).
|
|---|
| Figure 5. PETase (Hour 8). |

|
|---|
| Figure 6. LCC (Hour 8). |
|
|---|
|
|---|
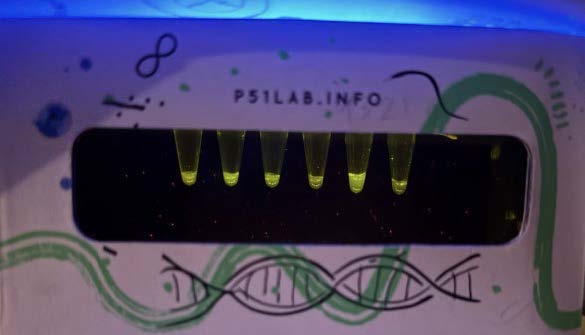
| Figure 7. 100ng/ul PETase at hour 18. |

|
|---|
| Figure 8. 100ng/ul LCC (original) at hour 18. |
|
|---|
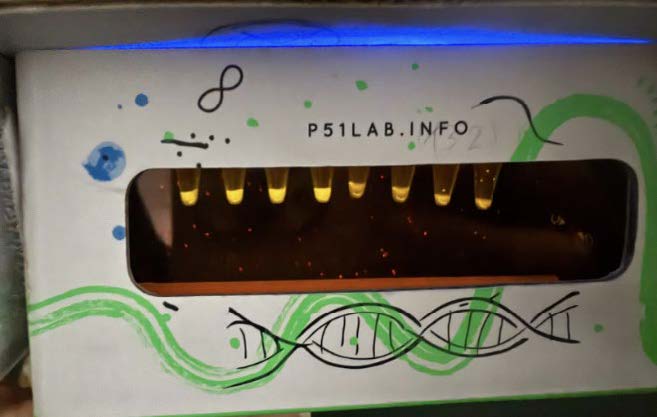
| Figure 9. 100ng/ul LCC (thermostable) at hour 18. |
|
|---|
| Figure 10. LCC Dried Out. |
|
|---|
Quantitative assessment of plastic powder circularity via ImageJ
Circularity of the plastic powder particles (250X) was quantified using ImageJ
|
|---|
| Figure 11. 100ng/ul LCC (original) at hour 18. |

|
|---|
| Figure 12. LCC (original) 250X. |

|
|---|
| Figure 13. LCC (modified) 250X. |

|
|---|
Discussions
The successful use of PETase and LCC provides a strategy for reducing the growing amount of plastic waste entering ecosystems each year. By enabling enzymatic degradation of PET, this project contributes to the goal of developing environmentally friendly methods for plastic recycling and waste reduction. However, the experiments encountered several challenges that may have affected the results and data collected.
Protein purification
The enzymes were not purified prior to the experiment, making it difficult to determine their exact concentration. The limitation of purifying the enzymes allowed for uncertainty regarding the quantity and functionality of the proteins during plastic degradation. In order to determine the effect of protein purification in this experiment, future experiments will contain chemically filtered proteins. There is a His-tag on the GFP within the plasmid constructs that would allow for this (Figures 1-3).
Cell-free systems don’t guarantee correct protein folding
Without verification methods such as Western blotting, there is no confirmation that functional PETase or LCC enzymes were synthesized. As a result, the presence of GFP fluorescence could not be directly linked to active enzyme production, and it remains unclear whether the intended proteins were actually properly formed. This means that differences in enzyme activity may truly be due to the enzymes themselves or as a consequence of cell-free expression and improper folding. With proper characterization, enzyme structure can be better understood.
Inconsistency with BioBits®
In the second round of experimentation, a mixture of new and older (1-year-old) BioBits® kits was used, which may have introduced variability in results. For example, PETase failed to express GFP in the second round when it did previously in the first experiment, possibly due to degradation of reagents or inconsistencies between BioBits® kits. In future experiments, all BioBits® kits should be new to optimize expression and reduce variability.
Inconsistency with the values of enzymes
Due to varying results and data from the experiments, some likely resulted from pipetting errors made during preparation for the examples. The difference in the amount of enzyme changes the concentration, and because it involved small values and non-purified enzymes, minor inaccuracies could have made significant variation in concentrations. Improved pipetting techniques could resolve this issue, but more precise and careful pipetting in future experiments will help ensure the concentrations remain as accurate as possible.
LCC dried out when incubated at 72°C
When LCC was incubated in the second experiment, some reactions appeared to dry out during this process. This can be due to the small volume of LCC added. Additionally, if drying altered the reaction conditions or damaged the enzyme, it could have limited its degradation efficiency. In the future, tubes should be wrapped with Parafilm to ensure that the enzyme does not dry out while incubating. Plastics will also be monitored in 72-degree reactions to ensure the LCC does not dry out.
Removing impurities from PET powders and films
While trying to remove the plastic powders and films from PETase and LCC by rinsing in nuclease-free water, there were likely contaminants on the PET film and powders, such as DNA or BioBits® ingredients. This did not seem to interfere with SEM imaging, but is a consideration for follow-up experiments.
Inability to measure differences in PET films
The assessment of PET film degradation proved difficult due to the smooth and uniform texture of the film. The flat surface lacked clear visual features that could be tracked over time, and the size of the film made it impossible to use the ImageJ circularity feature to detect significant change. In the following experiments, if the spots visible on the SEM scans are related to enzyme activity, the diameter of the spots can be measured.
Difficulty removing background in SEM scans of PET powders
It was difficult to quantitatively analyze the circularity in PET powders, as there were challenges in isolating the particles from the background in SEM scans. While the “Find Edges” and threshold functions in ImageJ were applied to enhance particle outlines, variations in contrast and lighting made it difficult to consistently differentiate particles from their background. This may have affected the accuracy of circularity measurements. The data gathered from the experiment using ImageJ to find the difference between the PET powders and the negative control shows the difference as “-0.002.” Because the average circularity was analyzed, the powders in the PETase sample could have been less circular compared to the sample of PET powders in the negative control.
Plasmid constructs with T7 promoter and T7 terminator
The T7 promoter and T7 terminator in the plasmid constructs (Figures 1-3) were needed for BioBits® to work. They are strong genetic parts that originated in bacteriophage viruses. Strong genetic parts are needed for high affinity to the cell-free ingredients, such as RNA polymerase. However, it should be noted that these constructs would be unable to hypothetically be used in cells as they are, because the T7 promoter is so strong that it would attract a cell’s RNA polymerase and other molecules away from any of the cell’s own transcription and translation. Therefore, if these specific constructs were attempted to be used in cells, the cells would likely not survive.
Next steps
In future experiments, it will be crucial to address the issues highlighted in the discussion section, such as maintaining consistency with DNA and BioBits®, as well as implementing additional precautions to prevent DNA dehydration during the incubation process. Moreover, further research should be performed to explore more effective methods to determine differences in before and after PET films and powders from SEM scans. Finally, experiments incorporating these strategies should be conducted to confirm or enhance the data collected.
Acknowledgements
The authors would like to acknowledge Dr. Lindsey L’Ecuyer for her support in our research and her guidance with lab experiments. We also wish to thank Andover Coalition for Education (ACE) for providing the necessary funding to carry out this project. We are very grateful for our mentors, Nathan Crook and Tianyu Li at NCSU, for guiding us with research and providing us with information and lab techniques necessary. Lastly, the authors thank all the original students, Puloma Bishnu and Krishnaveni Dole, as well as last year’s students for their contributions to the foundational work of this research.
Students from last year: Evelyn Wheel, Megan Zhang, Hannah Lehman, Miley Arora, Aarit Chauhan
References
Organisation for Economic Co-operation and Development. (2022, February 22). Plastic pollution is growing relentlessly as waste management and recycling fall short, says OECD. Retrieved April 9, 2025, from https://www.oecd.org/en/about/news/press-releases/2022/02/plastic-pollution-is-growing-relentlessly-as-waste-management-and-recycling-fall-short.html