Shreya Soni, Andover High School, Andover, MA
Reviewed on 3 May 2025; Accepted on 9 June 2025; Published on 27 October 2025
With help from the 2025 BioTreks Production Team.
Celiac disease is a serious autoimmune disorder that damages the small intestine in response to gluten, a protein found in wheat, rye, and barley. With an estimated 1 in 100 people worldwide affected and the prevalence continuing to rise, there is a growing need for innovative treatments that go beyond strict gluten-free diets. Current therapies rely heavily on dietary restriction, which can be difficult to maintain and do not always prevent inadvertent exposure. This project proposes a synthetically engineered Escherichia coli Nissle 1917 (EcN) probiotic designed to reduce gluten-induced inflammation in individuals with celiac disease. The engineered EcN strain detects elevated levels of tumor necrosis factor-alpha (TNF-α), a key pro-inflammatory cytokine upregulated during a gluten-triggered immune response. In response, it secretes interleukin-10 (IL-10), a potent anti-inflammatory cytokine, into the gut environment. By modulating the immune response at the source, this approach aims to reduce intestinal inflammation and improve quality of life for individuals with celiac disease. Incorporating advanced genetic engineering, biosensing, and safety mechanisms, this project explores a novel therapeutic strategy that directly targets the underlying inflammatory pathway of the disease.
Keywords: Celiac disease, Escherichiacoli, inflammation, TNF-α, IL-10
Authors are listed in alphabetical order. Lindsey L'Ecuyer mentored the group. Please direct all correspondence to lindsey.lecuyer@andoverma.us.





Background
Celiac disease (CD), also known as a gluten sensitive enteropathy, is an autoimmune inflammatory condition triggered by an immune-mediated response to dietary gluten (a protein found in wheat, barley, and rye) in genetically predisposed individuals (Mayo Clinic, 2023). Affecting approximately 1 in 100 people, CD is driven by an inappropriate immune response to gluten peptides, particularly in individuals obtaining the HLA-DQ2 or HLA-DQ8 genetic haplotypes (Posner & Haseeb, 2025). Upon gluten ingestion, the protein is broken down into peptides that the immune system recognizes, initiating the activation of both innate and adaptive immune responses (Ciccocioppo et al., 2005). Specifically, this response correlates to damage in the small intestinal mucosa—the three-layered membrane that lines the insides of the body and secretes mucus to capture invaders— leading to villous atrophy, gastrointestinal symptoms, malabsorption of important nutrients, and a range of symptoms including abdominal pain, diarrhea, dermatitis herpetiformis, osteoporosis, and neuropathy (Celiac Disease Foundation, 2025).
A distinctive attribute of CD is the inflammatory response in the small intestine that is crucial in the progression of the disease (Barone et al., 2022). Inflammation is the body’s response to illness, injury, or a foreign object such as germs or toxic chemicals (Cleveland Clinic, 2021). Usually, it is a protective mechanism, but in people with CD, it becomes dysregulated and instead leads to chronic inflammation and tissue damage (Barone et al., 2022). Pro-inflammatory cytokines, in particular tumor necrosis factor-alpha (TNF-α), are key factors in this process (Cleveland Clinic, 2023). TNF-α is produced predominantly by activated macrophages, T cells, and natural killer cells and exists in both soluble and transmembrane forms. It binds to TNFR1 and TNFR2 to trigger signaling cascades related to inflammation, cell survival, and immune cell requirement. TNF-α is a major aspect of many autoimmune diseases, including rheumatoid arthritis, inflammatory bowel disease, and celiac disease, due to its nature to exacerbate immune response, disrupt epithelial integrity, and promote fibroblast activation (Jang et al., 2021). Moreover, elevated TNF-α levels give rise to tissue damage and chronic inflammation in the celiac gut, making it a key therapeutic target in the management of CD (Rezaei-Tavirani et al., 2019).
While current management of CD is solely dependent on lifelong gluten-free diets, adherence is difficult, and cross-contact with gluten-enriched food is common. Cross-contact often occurs through shared kitchen equipment, restaurant meals, mislabeled products, or trace contamination during food process due to the lack of options available in the market and unregulated health practices in terms of food handling and labeling practices (Cross-Contact | BeyondCeliac.org, n.d.). Furthermore, there are no definitive cures or therapies available for CD that are FDA approved, with many treatments like Ritlecitinib by Pfizer (a kinase inhibitor that targets JAK3 and TEC-family tyrosine kinases, disrupting inflammatory and immune signaling pathways also implicated in alopecia areata, that is currently being researched as a potential cure for celiac disease) and Amlitelimab by Sanofi (a fully human, non-T cell depleting monoclonal antibody that blocks OX40-Ligand, aiming to reduce gluten-induced intestinal changes in patients with non-responsive celiac disease) still in the research and development phase (Celiac Disease Foundation, 2018). Thus, the necessity for novel treatments that address the inflammatory and intestinal homeostasis aspects of CD have never been greater.
One promising anti-inflammatory target is interleukin-10 (IL-10), a cytokine secreted by many cells such as macrophages, dendritic cells, and T cells. IL-10 inhibits pro-inflammatory cytokines like TNF-α by signaling through the JAK1-TYK2-STAT3 pathway, limiting damage to tissue and promoting immune tolerance. Most importantly, IL-10 supports the repair of the intestinal epithelial lining, an important facet in the recovery of patients with CD (Manavalan et al., 2010). Although IL-10 can exhibit pro-inflammatory effects under certain conditions, particularly in the presence of uncontrolled immune activation or in cancer microenvironments, its role in the gut in response to TNF-α is primarily anti-inflammatory and immunoregulatory. In this context, IL-10 functions to restore mucosal homeostasis and suppress excessive inflammation, making it a suitable therapeutic candidate for CD (Islam H et al. 2021).
To address the challenges of treating chronic intestinal inflammation in celiac disease, prior therapies have relied on anti-TNF-α monoclonal antibodies like infliximab and adalimumab, which block TNF-α or its receptor interactions. While these have shown success in other autoimmune diseases, they are often expensive, systemically immunosuppressive, and can have negative side effects such as carrying risks for infections and lymphoma (Hoey et al., 2019; Lee et al., 2024). Escherichia coli (E. Coli) is a safer and targeted alternative due to its delivery of therapy locally in the gut, which avoids systemic immunosuppression. As such, this research aims to engineer E. coli Nissle 1917 (EcN) to function as a localized and responsive anti-inflammatory system that triggers secretion of human IL-10 to dampen intestinal inflammation and restore immune homeostasis in patients with CD. To achieve this, EcN will be engineered with a plasmid incorporating a constitutively expressed Lpp-OmpA–anti-TNF-α nanobody fusion protein under the J23119 promoter for TNF-α detection, and a PcpxP stress-response promoter controlling the expression of a YebF-IL-10 fusion protein, enabling targeted secretion of IL-10 in response to inflammatory stress.
Systems level
The proposed biological system is a living therapeutic consisting of a chassis that will function autonomously within the gastrointestinal tract of individuals with CD. This system is built around a genetically engineered chassis that will operate using a synthetic stimulus-response circuit to detect inflammatory cues and deliver a localized therapeutic response. The main goal of this system is to detect TNF-α due to its role in immune activation and mucosal damage in CD. TNF-α is the input signal for the system that will prompt a set of genetic events once it has been recognized by the chassis (Figure 1). The sensing module features a surface-anchored nanobody that binds specifically to extracellular TNF-α, which starts the signal transduction process. This nanobody, derived from camelid heavy-chain-only antibodies, is the monovalent candidate VHH#2B, selected for its high affinity, stability, and >90% sequence homology to human VH regions, reducing immunogenicity and enabling efficient TNF-α detection in the gut (Muyldermans, 2013; Part:BBa K4387996 – Parts.igem.org, 2022). Furthermore, this interaction will start the downstream genetic pathway through a promoter responsive to the TNF-α binding induced membrane stress. The physical interaction between the chassis and TNF-α via the nanobody disturbs the neutrality of the bacterial outer membrane, triggering a stress response pathway within the cell. This stress response activates specific regulatory proteins that initiate transcription from a stress-responsive promoter.This activation will lead to the transcription and translation of the human IL-10 gene which will later dampen immune responses and promote epithelial healing in the gut (Saraiva et al., 2019). To ensure effective delivery of the IL-10, the gene will be fused to a secretion tag that facilitates its export from the chassis into the external environment, where it will be able to directly suppress inflammation, restore immune tolerance, and lessen the severity of the symptoms caused by CD. The genetic components that make up this series of events/circuit are carried on a modular plasmid that will be injected into a cell of the chassis which enables efficient construction, reproducibility, and adaptability. Additionally, this plasmid based design will allow the genetic components and chassis to be easily modified, making it possible to reprogram the chassis to detect and respond to TNF-α if the original plan is unsuccessful or needs adjustment based on patient-specific inflammatory profiles (Brophy & Voigt, 2014).
| Figure 1. Schematic representation of an engineered microbial chassis designed for therapeutic intervention in the gut. Upon detection of the pro-inflammatory cytokine TNF-α in the intestinal environment, the chassis responds by producing and secreting the anti-inflammatory cytokine IL-10. This system aims to mitigate inflammation and promote intestinal immune homeostasis. |
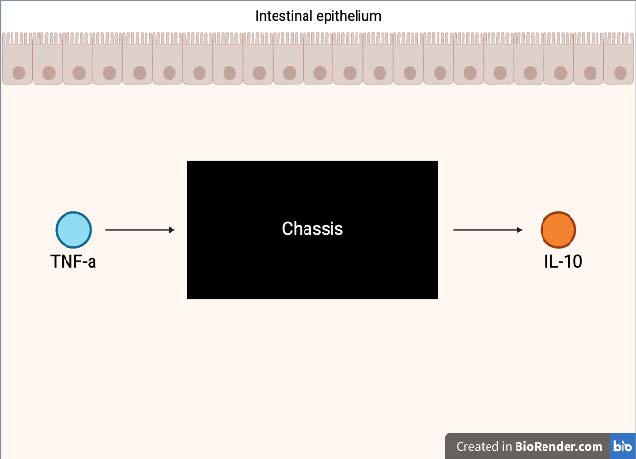
|
|---|
Device level
The proposed chassis for this system is EcN. Isolated by Professor Alfred Nissle from Freiburg, Germany from the intestinal microflora of a young soldier who didn’t develop infectious diarrhea during the Shigella outbreak in World War I, EcN is a gram-negative bacterium that has conventionally been used in many gastrointestinal disorders including diarrhea, uncomplicated diverticular disorder, and ulcerative colitis. This strain of E. coli lacks common virulence factors that make bacteria harmful such as a-hemolysis and P-fimbrial adhesins which makes it an optimal device. EcN contains microcins and multiple iron uptake systems along with adhesins and proteases which allows it to outcompete other microbes by efficiently gathering iron, adhere to the gut lining, and reinforce its ability to establish stable colonization within the gut microbiome. Additionally, EcN doesn’t produce any known toxins, is serum sensitive, and has a non-toxic lipopolysaccharide (LPS) structure, meaning that it is easily controlled by the body and doesn’t overstimulate the immune system (Scaldaferri et al., 2016). As can be seen, these characteristics make EcN an ideal candidate for the delivery of engineered probiotic therapies that must function safely and effectively within the human gastrointestinal tract.
The core functionality of the device is achieved through a genetically engineered EcN that carries a detection and response system based on TNF-α. The device is designed to detect TNF-α and respond by secreting IL-10 in order to modulate the inflammatory response associated with CD. Although studies have shown that EcN itself can induce gut cells to produce IL-10 within the presence of EcN, this approach will ensure that IL-10 is not unnecessarily produced in the absence of inflammation (Scaldaferri et al., 2016). By tightly linking cytokine detection to therapeutic response, the system minimizes the risk of immune disruption and enables precise, condition-specific delivery of IL-10 to inflamed intestinal tissue.
To enable this functionality, many key modules must be introduced into EcNs genetic architecture. This includes a sensing module, a signal transduction module, and response module that are detailed below.
Sensing module
As the initial component of the genetic circuit, the sensing module will be responsible for detecting TNF-α in the gut environment. In order to do this, a nanobody that specifically binds to TNF-α is expressed on the surface of EcN, meaning that the EcN will be genetically modified to have the nanobody on its surface. The nanobody will be fused to an outer membrane protein like Lpp-OmpA, which anchors the nanobody onto the surface of the bacterium and ensures that the nanobody directly interacts with the extracellular TNF-α in the gut (Part:BBa K1991004 – Parts.igem.org, 2016). This interaction occurs when the extracellular TNF-α that is released upon the consumption of gluten binds to the nanobody, initiating the process of TNF-α detection by the EcN. The surface expression of the nanobody allows the EcN to efficiently recognize and respond to the presence of TNF-α without internalization, simplifying and fortifying the detection process of the system (Lee et al., 2024).
Signal transduction module
Upon binding of the nanobody to TNF-α, the signal transduction module becomes activated and links the TNF-α binding to downstream gene expression. This means that the molecular recognition of TNF-α by the nanobodies on the EcN surface will be translated into a biological response within the EcN chassis. The pCpxP promoter, positioned upstream of the gene encoding the anti-inflammatory cytokine IL-10, acts as a sensor of envelope or membrane stress → conditions that worsen in the inflamed gut environment characteristic of celiac disease. When TNF-α binds to the surface displayed nanobody, the interaction induces localized stress at the outer membrane, activating the CpxAR two-component system (Zhang et al., 2005). Activated CpxR then promotes transcription from the pCpxP promoter, leading to expression of IL-10. This design ensures that the binding of TNF-α triggers a tightly controlled transcriptional response, enabling the engineered EcN to produce and secrete IL-10 precisely in response to inflammatory signals (Saraiva et al., 2019; Hoey et al., 2019).
Response module
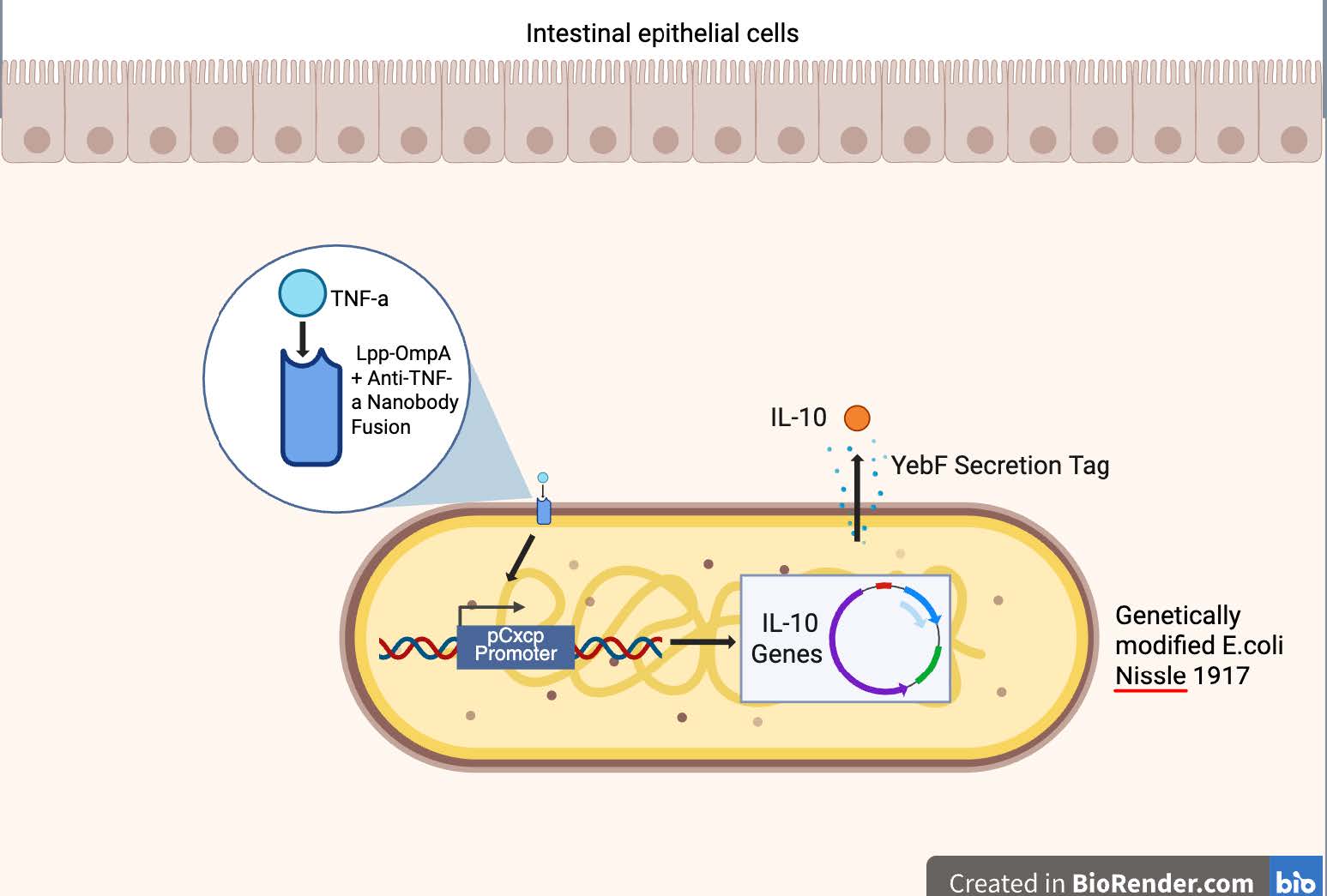
The final module of the circuit is the response module, and it manages the execution of therapeutic action triggered by the detection of TNF-α (Ciccocioppo et al., 2005). For our purposes, the output is the secretion of IL-10, which will reduce inflammation in the gut through the inhibition of pro-inflammatory cytokines like TNF-α (Figure 2). Upon the activation of the signal transduction module, EcN expresses L-10 coding sequence, which is then exported out of the bacterium using a secretion system like the YebF transporter protein. These secretion tags will cause the IL-10 to be released into the gut environment efficiently where it can interact with the gastrointestinal tract to reduce the effect of pro-inflammatory cytokines, alleviate CD symptoms, and most importantly, limit tissue and small intestine damage (Zhang et al., 2005).
| Figure 2. Schematic representation of the engineered E. coli Nissle 1917 therapeutic system for inflammatory regulation in celiac disease. The bacterium displays an Lpp-OmpA–anti-TNF-α nanobody fusion on its surface to detect the presence of TNF-α in the gut microenvironment. Upon binding TNF-α, the signal activates the pCpxP promoter, driving expression of IL-10 genes. The produced IL-10 is secreted via a YebF secretion tag, enabling local anti-inflammatory action near intestinal epithelial cells. |
|
|---|
Figure 2: Schematic representation of the engineered E. coli Nissle 1917 therapeutic system for inflammatory regulation in celiac disease. The bacterium displays an Lpp-OmpA–anti-TNF-α nanobody fusion on its surface to detect the presence of TNF-α in the gut microenvironment. Upon binding TNF-α, the signal activates the pCpxP promoter, driving expression of IL-10 genes. The produced IL-10 is secreted via a YebF secretion tag, enabling local anti-inflammatory action near intestinal epithelial cells.
Parts level
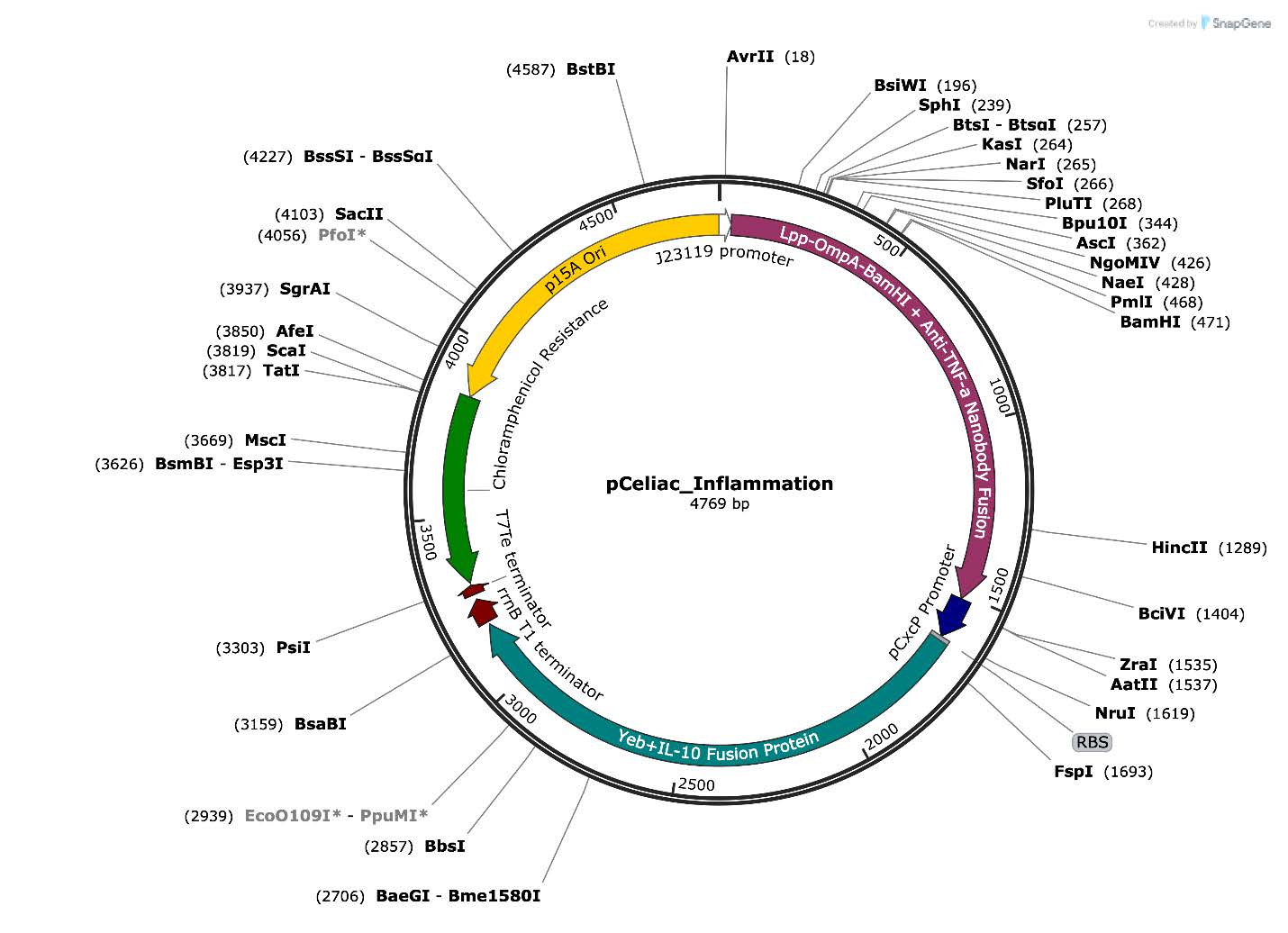
As mentioned previously, EcN will be the chassis that will induce IL-10 production upon TNF-α detection. Specifically, the plasmid designed for this system will be engineered to include all of the components necessary to carry out the expected outcome based on the expected input. Listed below are the parts along with their specific functions within the EcN (Table 1 and Figure 3).

|
|---|
| Figure 3. Engineered plasmid map encoding a TNF-α-responsive therapeutic system. This plasmid contains two expression cassettes: one featuring the constitutive J23119 promoter driving expression of a membrane-anchored Lpp-OmpA–anti-TNF-α nanobody fusion protein, and a second cassette with the inflammation-inducible PcpxP promoter driving secretion of an IL-10 anti-inflammatory cytokine fused to the YebF export signal. Standard elements include the B0034 ribosome binding site, B0015 double terminator, chloramphenicol resistance marker for bacterial selection, and the p15A origin of replication for plasmid maintenance in E. coli. |
|
|---|
Figure 4: Engineered plasmid map encoding a TNF-α-responsive therapeutic system. This plasmid contains two expression cassettes: one featuring the constitutive J23119 promoter driving expression of a membrane-anchored Lpp-OmpA–anti-TNF-α nanobody fusion protein, and a second cassette with the inflammation-inducible PcpxP promoter driving secretion of an IL-10 anti-inflammatory cytokine fused to the YebF export signal. Standard elements include the B0034 ribosome binding site, B0015 double terminator, chloramphenicol resistance marker for bacterial selection, and the p15A origin of replication for plasmid maintenance in E. coli.
Safety
The engineered EcN will be constructed and tested underneath Biosafety Level 1 as it is a defined organism and unlikely to cause disease (Trapotsis, 2022). Since it is classified at this level, this means that it is suitable to be handled by people obtaining different experience levels, ranging from seasoned industry professionals to high school students. Additionally, a strain of EcN obtains the GRAS (Generally Recognized As Safe) status under FDA approval, therefore decreasing any potential risk of harm (FDA, 2023). Biosafety mechanisms like kill switches will be integrated in order to ensure that the EcN will be confined to the gut and not travel to an external environment outside. This means that if the EcN experiences a change in oxygen, pH, temperature, or is in an environment with a lack of gut-specific nutrients, it will self-destruct (Sterling et al., 2017). Other safety measures that can be taken include adding more triggers to the EcN before it secretes IL-10, such as including other pro-inflammatory cytokines or adding a limiting gene that self-destructs after a certain amount of time or number of cell replications. These steps will ensure that the production of IL-10 occurs under stricter circumstances and ensure that the treatment is temporary, controlled, and renewable. Beyond this, containment measures like confining the EcN in petri dishes, using sterile techniques, and proper waste decontamination protocols will be applied throughout the project to prevent bacterial escape into the environment (LaVan & Marmon, 2010). All researchers involved in the execution portion of this research will be required to obtain training in biosafety practices and follow institutional guidelines for genetic engineering/synthetic biology. This will include the personal protective equipment like lab coats, gloves, and safety goggles, the proper sanitation of laboratory areas used during the experiments, as well as a strong foundation in handling genetically modified organisms in a responsible and careful fashion. As the project progresses, continuous risk assessments and regular updates to safety protocols will be performed to account for any modifications to the organism, system design, and laboratory/industry regulations (Trapotsis, 2022). These combined practices confirm the safe development, handling, and eventual testing of the engineered EcN, upholding both scientific integrity and public health standards.
Discussions
The engineered EcN presented in this project depicts a new and focused approach to addressing the chronic intestinal inflammation associated with CD. Through the design of the modular synthetic circuit with TNF-a detecting capabilities and localized excretion of IL-10, this therapy considers a key pathological driver of CD that is often overlooked, dysregulated immune response, while decreasing the systemic side effects that arise in conventional treatments (Lee et al., 2024; Saraiva et al., 2019). Additionally, the specificity of the TNF-α nanobody in this genetic circuit validates that the therapeutic response is solely based on active inflammation in the gut, streamlining the system and making it conditionally responsive. Compared to current clinical approaches towards CD like lifelong gluten-free diets and immunosuppressants, this probiotic-based system offers a non-invasive, sustainable, and cost-practical alternative (Celiac Disease Foundation, 2025; Islam et al., 2021). Using EcN as a chassis strengthens the durability and feasibility of the system because of its safety features, compatibility with the human intestine and gut microbiome, and ability to reproduce/clone quickly in comparison to other organisms (Scaldaferri et al., 2016; Trapotsis, 2022). Also, the IL-10 emission via the YebF lets therapeutic efforts take place without lysis of the EcN which preserves the functionality of the system (Jang et al., 2021). Nonetheless, several considerations remain. Since the inflammatory environment in CD patients varies in intensity and can be caused by a combination of cytokines, it may be difficult for the EcN system to recognize changes in TNF-α levels and significantly de-escalate inflammation (Hoey et al., 2019; Rezaei-Tavirani et al., 2019). Differences in gut microbiome composition, pH, and host genetics might influence the impact and colonization strategy of the engineered EcN, causing it to display a different behavior than intended. Challenges in scaling production, consistent plasmid retention, and the prevention of horizontal gene transfer of the engineered elements to native microbes also pose potential biosafety risks (Manavalan et al., 2010; Trapotsis, 2022). Taking all of this information into consideration, future research should continue to explore multi-cytokine detection circuits, the interactions between EcN and the gut, and refined biocontainment strategies. If correctly implemented, this system could revolutionize the way immune-mediated gastrointestinal diseases are treated through precision, biocompatibility, and patient-centered innovation.
Next steps
To further develop the system discussed in this project, many research steps can be taken within a high school synthetic biology lab setting.The initial focus will be on constructing the plasmid that contains the nanobody-based TNF-α sensor and IL-10 secretion module and includes gene synthesis/cloning of the nanobody sequence, fusion to a suitable membrane-bound/cytoplasmic signaling domain, and the confirmation of IL-10 activation in response to TNF-α (Cameron et al., 2014; Canton et al., 2008). Once the genetic circuit has been assembled, transformation of the plasmid into EcN or a lab-safe strain like E. coli DH5α should be performed. These experiments can include culturing the transformed cells and measuring fluorescence created from the ELISA-based detection of IL-10 in response to simulated inflammatory conditions which can be done by exposing the cells to cytokines or lipopolysaccharides (Mukherjee et al., 2021; Alper et al., 2005). The secretion system using the YebF carrier protein can also be tested by fusing IL-10 or a reporter like mCherry to YebF and assessing the presence of extracellular protein through confirmation methods like SDS-PAGE or Western blot. Safety and plasmid stability assay should also be conducted while advancing in this project by culturing engineered strains over several generations and evaluating the output of IL-10, plasmid presence through gel electrophoresis, and statistical data. Overall, following these steps will provide basic insights into the viability and practicality of this system by using simple conditions, equipment, and readily available material to refine the design for future computational modeling, in vivo, or clinical modeling (Canton et al., 2008; Mukherjee et al., 2021).
Author Contributions
SS was the sole author on this project and handled all components of it, utilizing the mentorship of Dr. Lindsey L’Ecuyer throughout the process
Acknowledgements
I would like to thank my BioBuilder club mentor, Dr. Lindsey L’Ecuyer, for her invaluable guidance and mentorship throughout the course of our project. Her expertise in the field of synthetic biology and her unwavering dedication to our success have been instrumental to the completion of this research. Furthermore, her passion for science and her enthusiasm for teaching have been an inspiration for all of us and have motivated us to pursue further research in the field of synthetic biology. I am truly thankful for her support and guidance, and we could not have completed this project without her.
References
Alper, H., Fischer, C., Nevoigt, E., & Stephanopoulos, G. (2005). Tuning genetic control through promoter engineering. Proceedings of the National Academy of Sciences, 102(36), 12678–12683. https://doi.org/10.1073/pnas.0504604102
Barone MV, Auricchio R, Nanayakkara M, Greco L, Troncone R, Auricchio S. Pivotal Role of Inflammation in Celiac Disease. Int J Mol Sci. 2022 Jun 28;23(13):7177. doi: 10.3390/ijms23137177. PMID: 35806180; PMCID: PMC9266393.
Brophy, J. A. N., & Voigt, C. A. (2014). Principles of genetic circuit design. Nature Methods, 11(5), 508–520. https://doi.org/10.1038/nmeth.2926
Cameron, D. E., Bashor, C. J., & Collins, J. J. (2014). A brief history of synthetic biology. Nature Reviews Microbiology, 12(5), 381–390. https://doi.org/10.1038/nrmicro3239
Canton, B., Labno, A., & Endy, D. (2008). Refinement and standardization of synthetic biological parts and devices. Nature Biotechnology, 26(7), 787–793. https://doi.org/10.1038/nbt1413
Celiac Disease Foundation. (2018). Future Therapies | Celiac Disease Foundation. Celiac Disease Foundation; Celiac. https://celiac.org/about-celiac-disease/future-therapies-for-celiac-disease/
Celiac Disease Foundation. (2025). What Is Celiac Disease? Celiac Disease Foundation. https://celiac.org/about-celiac-disease/what-is-celiac-disease/
Ciccocioppo, R., Di Sabatino, A., & Corazza, G. R. (2005). The immune recognition of gluten in coeliac disease. Clinical and Experimental Immunology, 140(3), 408–416. https://doi.org/10.1111/j.1365-2249.2005.02783.x
Cleveland Clinic. (2021). Inflammation: What is it, causes, symptoms & treatment. Cleveland Clinic. https://my.clevelandclinic.org/health/symptoms/21660-inflammation
Cleveland Clinic. (2023, March 1). What Are Cytokines? Types and Function. Cleveland Clinic. https://my.clevelandclinic.org/health/body/24585-cytokines
Cross-Contact | BeyondCeliac.org. (n.d.). Beyond Celiac. https://www.beyondceliac.org/gluten-free-diet/cross-contact/
ELISA Technique. (2023, May 15). Cleveland Clinic; Cleveland Clinic. https://my.clevelandclinic.org/health/articles/24990-elisa
FDA. (2023, October 17). Generally Recognized as Safe (GRAS). U.S. Food and Drug Administration. https://www.fda.gov/food/food-ingredients-packaging/generally-recognized-safe-gras
Hoey, R. J., Eom, H., & Horn, J. R. (2019). Structure and development of single domain antibodies as modules for therapeutics and diagnostics. Experimental Biology and Medicine, 244(17), 1568–1576. https://doi.org/10.1177/1535370219881129
Islam H, Chamberlain TC, Mui AL and Little JP (2021). Elevated Interleukin-10 Levels in COVID-19: Potentiation of Pro-Inflammatory Responses or Impaired Anti-Inflammatory Action? Front. Immunol. 12:677008. doi: 10.3389/fimmu.2021.677008
Jang, D., Lee, A-Hyeon., Shin, H.-Y., Song, H.-R., Park, J.-H., Kang, T.-B., Lee, S.-R., & Yang, S.-H. (2021). The Role of Tumor Necrosis Factor Alpha (TNF-α) in Autoimmune Disease and Current TNF-α Inhibitors in Therapeutics. International Journal of Molecular Sciences, 22(5). https://doi.org/10.3390/ijms22052719
LaVan DA, Marmon LM. Safe and effective synthetic biology. Nat Biotechnol. 2010 Oct;28(10):1010-2. doi: 10.1038/nbt1010-1010. PMID: 20944581; PMCID: PMC2956601.
Lee, N. J., Jung, M., Yang, H. Y., & Shim, H. (2024). A single-domain antibody library based on a stability-engineered human VH3 scaffold. Scientific Reports, 14(1). https://doi.org/10.1038/s41598-024-68680-5
Manavalan, J. S., Hernandez, L., Shah, J. G., Konikkara, J., Naiyer, A. J., Lee, A. R., Ciaccio, E., Minaya, M. T., Green, P. H. R., & Bhagat, G. (2010). Serum cytokine elevations in celiac disease: Association with disease presentation. Human Immunology, 71(1), 50–57. https://doi.org/10.1016/j.humimm.2009.09.351
Mayo Clinic. (2023, September 12). Celiac Disease. Mayo Clinic; Mayo Clinic Staff. https://www.mayoclinic.org/diseases-conditions/celiac-disease/symptoms-causes/syc-20352220
Mukherjee, S., Bassler, B. L., & Bassler, B. L. (2021). Bacterial quorum sensing in complex and dynamically changing environments. Nature Reviews Microbiology, 19(6), 371–384. https://doi.org/10.1038/s41579-021-00536-5
Muyldermans, S. (2013). Nanobodies: Natural single-domain antibodies. Annual Review of Biochemistry, 82, 775–797. https://doi.org/10.1146/annurev-biochem-063011-092449
Part:BBa B0034 – parts.igem.org. (n.d.). Parts.igem.org. https://parts.igem.org/Part:BBa_B0034
Part:BBa J23119 – parts.igem.org. (2020). Igem.org. https://parts.igem.org/Part:BBa_J23119
Part:BBa K082010 – parts.igem.org. (2025). Igem.org. https://parts.igem.org/Part:BBa_K082010
Part:BBa K554004 – parts.igem.org. (n.d.). Parts.igem.org. https://parts.igem.org/Part:BBa_K554004
Part:BBa K1991004 – parts.igem.org. (2016). Igem.org. https://parts.igem.org/Part:BBa_K1991004
Part:BBa K2026001 – parts.igem.org. (2016). Igem.org. https://parts.igem.org/Part:BBa_K2026001
Part:BBa K2560046 – parts.igem.org. (2018). Igem.org. https://parts.igem.org/Part:BBa_K2560046
Part:BBa K2817009 – parts.igem.org. (2018). Igem.org. https://parts.igem.org/Part:BBa_K2817009
Part:BBa K2926077 – parts.igem.org. (2019). Igem.org. https://parts.igem.org/Part:BBa_K2926077
Part:BBa K4387996 – parts.igem.org. (2022). Igem.org. https://parts.igem.org/Part:BBa_K4387996
Posner, E. B., & Haseeb, M. (2025, February 4). Celiac Disease. PubMed; StatPearls Publishing. https://www.ncbi.nlm.nih.gov/books/NBK441900/
Rezaei-Tavirani, S., Rostami-Nejad, M., Reza Vafaee, Ensieh Khalkhal, Aliasghar Keramatinia, Ehsani-Ardakani, M. J., & Mohammadreza Razzaghi. (2019). Introducing tumor necrosis factor as a prominent player in celiac disease and type 1 diabetes mellitus. Gastroenterology and Hepatology from Bed to Bench, 12(Suppl1), S123. https://pmc.ncbi.nlm.nih.gov/articles/PMC7011075/
Saraiva, M., Vieira, P., & O’Garra, A. (2019). Biology and therapeutic potential of interleukin-10. The Journal of Experimental Medicine, 217(1), e20190418. https://doi.org/10.1084/jem.20190418
Scaldaferri, F., Gerardi, V., Mangiola, F., Lopetuso, L. R., Pizzoferrato, M., Petito, V., Papa, A., Stojanovic, J., Poscia, A., Cammarota, G., & Gasbarrini, A. (2016). Role and mechanisms of action of Escherichia coli Nissle 1917 in the maintenance of remission in ulcerative colitis patients: An update. World Journal of Gastroenterology, 22(24), 5505. https://doi.org/10.3748/wjg.v22.i24.5505
Shetty, R. (2003, July 17). Part:BBa B0015 – parts.igem.org. Parts.igem.org. https://parts.igem.org/Part:BBa_B0015
Trapotsis, A. (2022, March 11). Biosafety Levels 1, 2, 3 & 4 | What’s The Difference? Consolidated Sterilizer Systems. https://consteril.com/biosafety-levels-difference/Zhang, G., Brokx, S., & Weiner, J. H. (2005). Extracellular accumulation of recombinant proteins fused to the carrier protein YebF in Escherichia coli. Nature Biotechnology, 24(1), 100–104. https://doi.org/10.1038/nbt1174