Tucker Marshall, Regina Rubiano, and Hayden Walmsley, Western Reserve Academy, Hudson, Ohio, United States
Reviewed on 3 May 2025; Accepted on 9 June 2025; Published on 27 October 2025
With help from the 2025 BioTreks Production Team.
Coral reefs are crucial to marine biodiversity, supporting approximately 25% of marine species and a $2.7 trillion global economy, including fishing and tourism. However, coral bleaching, driven by rising ocean temperatures, threatens ocean ecosystems. Bleaching occurs when thermal stress forces coral to expel their symbiotic algae, Symbiodinium (zooxanthellae). These algae are crucial for coral survival as they provide energy, disease protection, and vibrant coloration, critical for photosynthesis and UV protection. Without these algae, corals become vulnerable to disease and death. As of 2024, 77% of corals have experienced bleaching-level heat stress, with temperatures projected to rise by 2.4°C by 2050. Current strategies, such as reef-safe sunscreens, carbon-sequestration seagrass, and emission cuts, offer localized relief but don’t tackle coral heat resistance directly. Our design aims to boost coral heat tolerance by transferring heat shock proteins (HSPs) from Arabidopsis thaliana into Symbiodinium using a gene gun. HSPs are proteins that maintain cellular homeostasis under stress, protecting against thermal damage by folding and repairing damaged proteins, thus preventing cellular dysfunction. A. thaliana exhibits advanced HSP mechanisms, including high oxidative stress management and protein repair, making its HSPs ideal candidates for boosting Symbiodinium heat resistance. Our approach offers a scalable method to increase coral resilience to rising ocean temperatures by strengthening the coral at a cellular level. By complementing global efforts, our solution strengthens coral biology by enhancing heat resistance mechanisms. This approach reduces thermal stress on Symbiodinium and promotes long-term coral survival, boosting marine ecosystem health and the global economies reliant on healthy reefs.
Keywords: Arabidopsis thaliana, Symbiodinium, heat shock proteins, HSP20, coral bleaching
Authors are listed in alphabetical order. mentored the group. Please direct all correspondence to .






Background
Coral reefs are among the most vital ecosystems in the ocean, providing shelter and enabling vast biodiversity across the globe. Although they only cover less than one percent of the seafloor, corals support at least 25% of marine species by providing protection and sustenance (United Nations Environment Programme [UNEP], 2025). Coral reefs also provide a valuable economy to humans, contributing to coastal protection and making up a cumulative 2.7 trillion dollar industry that includes fishing, restaurants, medical products, and more. Corals are marine invertebrates that belong to the phylum Cnidaria (NOAA Coral Reef Information System, 2025.). Corals are composed of polyps, a small tube structure with stinging tentacles used to catch prey and protect them from predators. Corals can be classified into two main types: soft and hard. Examples of soft coral include sea fans and sea feathers, whereas hard corals are the reef builders (Great Barrier Reef Foundation, 2023). Hard corals extract calcium and carbonates from seawater to build their outer skeletons made of limestone (National Geographic Society, 2024). The continuous building of this limestone is what creates coral reef ecosystems. Figure 1 shows an example of healthy coral as seen in Palmyra Atoll National Wildlife Refuge.
| Figure 1 Healthy Coral at Palmyra Atoll National Wildlife Refuge. (“Coral Reef at Palmyra Atoll National Wildlife Refuge” by USFWS Headquarters is licensed under CC BY 2.0.) |

|
| Figure 2 Coral Bleaching in the Gulf of Thailand After Heat Shock (“Coral bleaching in the Gulf of Thailand_11” by Eco Cafe Pranburi is licensed under CC BY-SA 2.0.) |

|
Impacts of coral bleaching
As of October 2024, it is estimated that approximately 77% of the world’s coral reef areas have experienced bleaching-level heat stress (Withers & Dickie, 2024). Furthermore, between 2009 and 2018, there was a 14% decline in global coral cover, mostly due to bleaching events (UNEP, 2025). This dramatic loss of coral has caused significant economic damage to industries such as fishing, tourism, restaurants, and the biomedical sector, while also costing governments hundreds of millions of dollars. For example, in Florida, projections show that bleaching could increase the coastal flood risk to more than 7,300 people and cost up to 823.6 million dollars annually (Coastal and Marine Hazards and Resources Program, 2023). Coral bleaching continues to occur due to rising temperatures. One model shows temperatures could increase by 2.4 degrees Fahrenheit by 2050 (Lindsey & Dahlman, 2024). This continuous and dramatic increase in temperatures will continue to harm economies and marine ecosystems.
| Figure 3. Seagrasses: Schools of fish swim around kelp forests for protection and food. (“15166-fish swarm through the kelp forest” by oliver.dodd is licensed under CC BY 2.0.) |

|
Current solutions for coral bleaching
Additionally, researchers have found that seagrass beds and kelp forests play a crucial role in reducing coral bleaching. A six-year study by University of California Davis professors and students found that seagrasses such as those seen in Figure 3 can reduce local acidity by up to 30 percent (Kerlin, 2021). According to the Monterey Bay Aquarium (n.d.), seagrasses absorb carbon dioxide from the ocean water when they photosynthesize, which raises the pH to higher levels , helping mitigate some of the effects of ocean acidification. Kelp forests may provide a buffer for sensitive animals (Monterey Bay Aquarium, n.d.), indirectly supporting biodiversity and creating an environment where coral can flourish. These carbon sequestration methods are natural and help lower localized ocean temperatures; however, extensive kelp declines in recent years (Greater Farallones National Marine Sanctuary, n.d.) due to overgrazing have left scientists stumped on what to do next.
Heat shock proteins
Both prokaryotic and eukaryotic cells produce HSPs, a group of protective proteins, in response to stressful conditions like temperature spikes (Wang et al., 2020). These proteins assist in folding, assembling, and disassembling other proteins, regulate protein degradation, facilitate protein movement across membranes, and inhibit cell death pathways as shown in Figure 4. HSPs play a crucial role in coral’s cytoprotective mechanisms, making them vital for resilience under thermal stress (Voolstra & Ziegler, 2020). However, prolonged heat exposure gradually reduces coral HSP expression levels, diminishing their protective effects. Since rising ocean temperatures are the greatest threat to coral reefs, introducing HSPs to Symbiodinium could enhance coral heat resistance and improve reef survival.
| Figure 4. Function of HSPs in stress response (Heat Shock, n.d.): The HSPs stabilize and refold proteins under stress to maintain cellular function. |
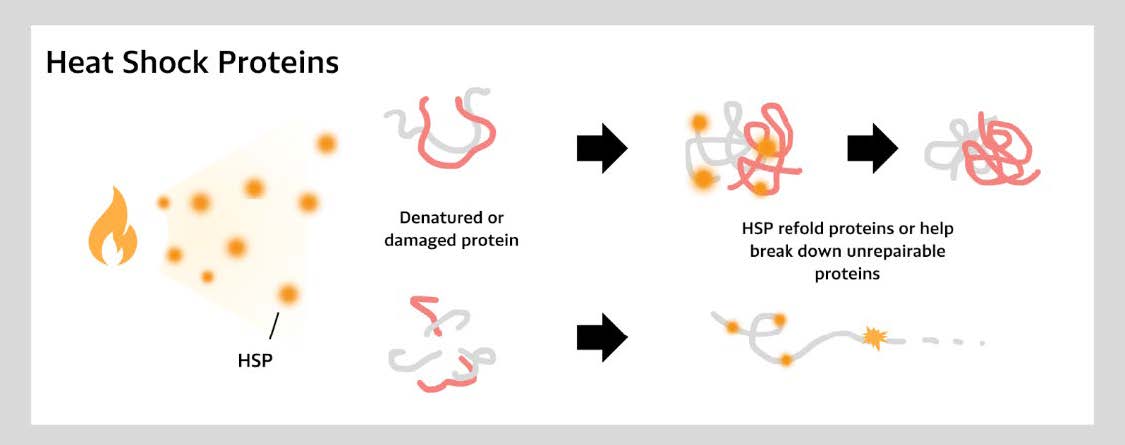
|
Our project aims to enhance the effectiveness of HSPs in Symbiodinium during extended periods of elevated temperatures. Given their exceptional stress tolerance properties, we selected HSPs from Arabidopsis thaliana (Figure 5) as promising candidates for strengthening heat resistance in Symbiodinium. Arabidopsis plants respond to prolonged heat with various adaptive strategies (Wang et al., 2020). These include increased transpiration, where plants lose water vapor to regulate temperature. Prolonged heat exposure produces reactive oxygen species (ROS), which are oxygen-containing molecules that act as signaling agents and can be sources of cellular damage. To counteract this, A. thaliana induces antioxidant enzymes to neutralize ROS and protect cells during stress (Wang et al., 2020). Finally, Arabidopsis activates the HSFA1 heat stress response pathway, where the transcription factor HSFA1 regulates the expression of HSPs that protect and repair heat-damaged proteins (Wang et al., 2020). In plants, transpiration cools the leaf surface. In coral, a similar adaptation could regulate surface temperatures or improve nutrient flow, creating a buffer against thermal stress. Enhanced electron transfer in response to heat stress could improve coral symbionts’ ability to maintain energy production under stress, stabilizing the coral’s internal environment during warm periods (Levin et al., 2017). Inducing a controlled ROS response and antioxidant enzymes could help corals better manage oxidative stress during heat events, reducing cellular damage (Wang et al., 2020).
| Figure 5: Arabidopsis thaliana: Also known as Thale Cress, a plant renowned for its stress tolerance. (“Arabidopsis thaliana inflorescencias” is licensed under CC BY-SA 3.0.) |

|
While these mechanisms are inherent to HSPs across various species, Arabidopsis HSPs stand out due to their stronger capacity for stress tolerance, making them ideal for strengthening coral survival under rising ocean temperatures.
Benefits of HSPs
Current and proposed solutions to coral bleaching primarily focus on mitigating ocean warming through reduced emissions or local cooling strategies, without directly addressing the coral itself. While reducing emissions is essential to address climate change, these approaches alone have limited short-term effects on rising ocean temperatures and don’t provide immediate support for coral reefs facing stress (Montalbetti et al., 2021), unlike a direct change to Symbiodinium heat regulation. Local cooling strategies, such as shading reefs or using artificial upwelling, are temporary and logistically challenging to scale globally, especially given coral ecosystems’ vast and widespread nature.
Our project offers a more sustainable and scalable solution by focusing on coral biology. By introducing HSPs from A. thaliana into Symbiodinium, we aim to directly enhance the coral’s natural heat tolerance at a cellular level. This approach addresses the effects of thermal stress and strengthens the coral’s intrinsic ability to survive in warmer conditions over time (Levin et al., 2017). Enhancing cellular resilience within coral symbionts could allow reefs to adapt naturally to their changing environment, providing a buffer against future thermal stress events without reliance on extensive external intervention (Levin et al., 2017). This method leverages the coral’s survival mechanisms, making it an innovative, targeted, and potentially long-lasting solution to the coral reef crisis.
| Figure 6. Symbiodinium through a Microscope: Symbiodinium and coral have a symbiotic relationship in which the algae live within the coral on a microscopic level. https://www.psu.edu/news/research/story/widespread-coral-algae-sym |

|
Systems level
Symbiodinium is essential to the health of polyps and ultimately coral reefs, sharing a symbiotic relationship as seen in Figure 6. For this reason, HSPs from A. thaliana have been selected to modify Symbiodinium (Swindell et al., 2007). Specifically, HSP20 was chosen due to its effectiveness in heat resistance and oxidative stress (Cui et al.,, 2021). Other HPSs, such as HSP-70 and HSP-90, are less reactive to heat and tend to have lower oxidative stress resistance. To integrate HSP20 into Symbiodinium, we employed a gene gun to modify the Symbiodinium. This method was chosen because the gene gun allows for the direct delivery of DNA into cells, bypassing the need for chemical or viral transfection methods (Carnegie Mellon University Environmental Health & Safety, 2024).. The new system involves a group of modified Symbiodinium existing within the walls of coral polyps. The modified Symbiodinium now possesses the ability to fold proteins in higher ocean temperatures, and if oxidative stress is present. The Symbiodinium assists in regulating protein folding within the coral polyp (Chen et al., 2019). As Symbiodinium better regulates its protein folding process,it should theoretically bolster the coral polyps’ protein folding. This creates a feedback loop of sustained protein stability in both symbiont and host. Our system should, then, be able to withstand increased ocean temperatures.
Device level
To enhance coral resilience against thermal stress, we propose introducing HSP20 genes from A. thaliana into Symbiodinium, the symbiotic algae essential for coral survival. The primary challenge in genetically modifying Symbiodinium is in its permanently condensed nuclear chromosomes, which limit access to stable genomic integration (Team: GA State SW Jiaotong, 2020). To overcome this, we will target the DNA-containing chloroplasts of Symbiodinium using a ballistic gene particle delivery system (Figure 7).
| Figure 7. Method of Delivering DNA Using a Gene Gun Note. A gene gun delivers DNA into cells by shooting particles coated with genetic material at a high velocity. https://commons.wikimedia.org/w/index.php?curid=40535573 |
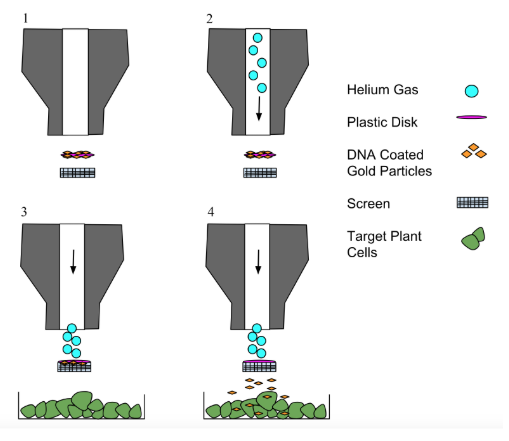
|
This method uses gold or tungsten microprojectiles coated with the plasmid DNA to deliver genes into Symbiodinium cells, targeting the chloroplast genome for stable expression (Kikkert et al., 2005). This approach allows for efficient gene expression without requiring nuclear genome modification, making it an efficient method for integrating HSP20.
The genetic construct includes a temperature-responsive eukaryotic promoter, specifically the Lambda repressor cI857 system, which induces the expression of the HSP20 gene when ocean temperatures exceed 30°C. The HSP20 gene encodes a small heat shock protein that enhances protein folding and increases oxidative stress resistance, aiding in cellular protection under thermal stress. To ensure proper transcriptional regulation, the construct incorporates the NOS terminator, which contains the dinoflagellate polyadenylation signal (AAAAG/C), facilitating efficient transgenic protein expression in Symbiodinium. (Team: GA State SW Jiaotong, 2020). Once introduced into Symbiodinium, the engineered algae will reside within coral polyps, providing thermal protection. The modified Symbiodinium will then show enhance protein-folding capabilities under thermal stress, potentially stabilizing the coral system.
Parts level
The genetic construct includes a temperature-responsive eukaryotic promoter, specifically the Lambda repressor cI857 system, which induces expression of the HSP20 gene when ocean temperatures exceed 30°C (Jechlinger et al., 1999). The HSP20 gene encodes a small heat shock protein that enhances protein folding, prevents protein misfolding, and increases oxidative stress resistance, thus aiding in cellular protection under thermal stress. To ensure proper transcriptional regulation, the construct incorporates the NOS terminator, which contains the dinoflagellate polyadenylation signal (AAAAG/C), facilitating efficient transgenic protein expression in Symbiodinium. (Team: GA State SW Jiaotong, 2020).
To ensure efficient translation in Symbiodinium, we will include an optimized ribosome binding site (Team: GA State SW Jiaotong, 2020). The genetic construct will be carried on a plasmid backbone containing replication and selection markers necessary for plasmid maintenance. Since Symbiodinium has permanently condensed nuclear chromosomes, stable transformation of its chloroplast genome is the most feasible approach. Together, these components form a tightly regulated system where Symbiodinium upregulates HSP20 in response to rising ocean temperatures, enhancing the coral’s ability to withstand thermal stress.
Safety
Symbiodinium is a BSL-1 organism and is thus considered very safe for laboratory experiments (Voolstra et al., 2021).We will utilize a heat-activated promoter toreduce the risk of overexpressing HSP20, increasing the viability of the modified Symbiodinium. Ironically, overexpression of HSP20 can interfere with typical protein homeostasis, boosting the possibility of misfolded proteins (Haghighi et al., 2014). Since Symbiodinium already exists in a natural symbiotic relationship with coral, using it as our chassis ensures compatibility with the host and avoids introducing a novel organism into the marine environment (Chen et al.,, 2019). However, genetic modification still presents potential risks. The stable transformation of chloroplast genomes helps minimize horizontal gene transfer, but we acknowledge that introducing engineered algae into open ecosystems must be approached cautiously. Furthermore, gene gun SOPs and safety procedures will be followed, and only people with Biological Safety Training will handle the gene gun (Carnegie Mellon University Environmental Health & Safety, 2024).
For experimentation, we will create our own temperature-regulated, small marine ecosystem with coral. This setup will include coral polyps and both unedited and edited Symbiodinium, allowing us to monitor algal colonization, coral health, and stress responses to elevated temperatures. Only after extensive testing and consistent results would we consider releasing the modified algae into the open ocean, ensuring the long-term stability and safety of the engineered Symbiodinium.
Discussions
As climate change continues to intensify, coral reefs are experiencing increasingly frequent and severe bleaching events. Our project proposes a novel solution for coral reef preservation by engineering thermotolerant Symbiodinium to better tolerate heat stress. By introducing the A. thaliana HSP20 into the chloroplasts of Symbiodinium, we aim to enhance the algal symbiont’s protein folding capacity and oxidative stress tolerance under thermal stress, thereby stabilizing the coral-algal symbiosis during ocean temperature spikes and prolonging coral life
The use of a temperature-responsive expression system, specifically the lambda repressor cl857 promoter, enables the conditional expression of HSP20 when ocean temperatures exceed 30℃,, a critical bleaching threshold for many coral species. This tunable control minimizes the metabolic burden on Symbiodinium under normal conditions while activating protective functions during heat stress, offering a targeted and efficient response to climate-induced bleaching events.
Targeting the chloroplast genome rather than the nuclear genome circumvents the challenge posed by the permanently condensed chromosomes in Symbiodinium, which have historically hindered stable transformation. Our biolistic transformation method offers a direct route to chloroplast integration, building on previous successes in algal systems. Using chloroplast-compatible regulatory elements, including the NOS terminators, provides flexibility in construct design. Notably, the NOS terminator contains the dinoflagellate polyadenylation signal (AAAAG/C), which has been associated with successful transgene expression in Symbiodinium through both silicon-carbide whisker and Agrobacterium-mediated transformation, suggesting compatibility within our system (Team: GA State SW Jiaotong, 2020).
If successfully expressed, HSP20 will assist Symbiodinium in maintaining proteostasis under thermal stress. This could create a positive feedback loop in which improved algal protein folding supports healthier coral hosts, stabilizing the symbiosis. While initial integration and expression studies will focus on in vitro assays, future experiments will assess the viability and functionality of modified Symbiodinium within coral polyps under heat-stress conditions. We would specifically be looking for increased Symbiodinium retention in coral during heat waves in the ocean and more tolerance to oxidative stress.
Next steps
Moving forward, we intend to perform the experiment within simulated coral environments to test the effectiveness of our modified Symbiodinium. These controlled settings will allow us to modify multiple variables such as elevated temperatures, changes in light intensity, and fluctuations in pH. We will test these metrics with the different clades of Symbiodinium. Different clades include Symbiodinium A, B, C, D, and each clade varies in heat tolerance and host compatibility (Chen et al., 2019). We plan to create eight simulated coral environments, to allow testing of four clades with our modifications and four control clades. To test the effectiveness of our genetic modification, we will monitor three primary markers, each with associated sub-indicators. These markers include 1. heat tolerance: cell viability (%), HSP20 expression levels, and reactive oxygen species (ROS) levels, 2. Symbiodinium health and productivity: cell density and photosynthetic efficiency (Fv/Fm), 3. Symbiodinium-coral symbiosis: host polyp health, glucose transfer to host (radiolabeled carbon tracking), and coral immune system response (ELISA or qPCR).
In addition to collecting vast data on the performance of our genetic modification, we also plan to collect data on the effectiveness of our promoter. Since timing is key in our construct, we will collect data on the conditions that activate the promoter. Conditions may include water temperature, oxidative stress, or UV exposure, and may unintentionally include non-stress conditions like mild temperature changes or light stress. The ultimate goal of promoter optimization is to ensure HSP20 is only expressed when necessary, minimizing unnecessary protein production, reducing metabolic burden, and preserving the overall health of our modified Symbiodinium.
Finally, we plan to complete a preliminary analysis of the ecological risks and consult with professionals such as marine scientists, biologists, synthetic biology experts, and environmental regulators. Conversations with these professionals will help guide the long-term plans of our design, ensure the safety of our applications, and promote ecological responsibility. With these detailed next steps, we hope to one day integrate our modified Symbiodinium into coral ecosystems in nature.
Author Contributions
All authors contributed equally to the writing of the Introduction. TM was responsible for the safety considerations and systems integration. RR led the development of the device and authored the Discussion section. HW contributed to the selection and assembly of parts, created the project video, and collaborated with TM to outline the next steps.
Acknowledgements
We would like to give a special thanks to Dr. Pethel for guiding us throughout this entire process. From laying out the entire project to proofreading our entire paper, Dr. Pethel has made this entire paper possible. We would also like to thank Dr. Leah Davis (NOAA) for her assistance on the technical side of our project. Dr. Davis made us think of our project in a new, unique light, making this a stronger overall paper.
References
Carnegie Mellon University Environmental Health & Safety. (2024). Standard operating procedure (SOP) for working safely with gene guns. Carneige Mellon University. https://www.cmu.edu/ehs/Laboratory-Safety/biological-safety/documents/ehs-standard-operating-procedures-for-working-safely-with-gene-guns1.pdf
Chen, J. E., Barbrook, A. C., Cui, G., Howe, C. J., & Aranda, M. (2019). The genetic intractability of Symbiodinium microadriaticum to standard algal transformation methods. PloS one, 14(2), e0211936. https://doi.org/10.1371/journal.pone.0211936
Coastal and Marine Hazards and Resources Program. (2023, August 9). Coral bleaching event can increase flood risk, economic losses. United States Geological Survey.. Retrieved November 14, 2024, from https://www.usgs.gov/programs/cmhrp/news/coral-bleaching-event-can-increase-flood-risk-economic-losses#:~:text=The%20USGS%20and%20partners%20have,(2023)%20coral%20bleaching%20event
Cui, F., Taier, G., Wang, X., & Wang, K. (2021). Genome-Wide Analysis of the HSP20 Gene Family and Expression Patterns of HSP20 Genes in Response to Abiotic Stresses in Cynodon transvaalensis. Frontiers in genetics, 12, 732812. https://doi.org/10.3389/fgene.2021.732812
Great Barrier Reef Foundation. (2023, April 24). What is coral? Retrieved November 22, 2024. https://www.barrierreef.org/news/explainers/what-is-coral
Greater Farallones National Marine Sanctuary. (n.d.). Kelp in Decline. https://farallones.noaa.gov/eco/kelp/decline.html
Haghighi, K., Bidwell, P., & Kranias, E. G. (2014). Phospholamban interactome in cardiac contractility and survival: A new vision of an old friend. Journal of Molecular and Cellular Cardiology, 77. https://doi.org/10.1016/j.yjmcc.2014.10.005
Hancock, L. (2025). Everything you need to know about coral bleaching—And how we can stop it. World Wildlife Fund. https://www.worldwildlife.org/pages/everything-you-need-to-know-about-coral-bleaching-and-how-we-can-stop-it
Jechlinger, W., Szostak, M. P., Witte, A., & Lubitz, W. (1999). Altered temperature induction sensitivity of the lambda pR/cI857 system for controlled gene E expression in Escherichia coli. FEMS microbiology letters, 173(2), 347–352. https://doi.org/10.1111/j.1574-6968.1999.tb13524.x
Jordan, R. (2022, May 6). Understanding how sunscreens damage coral. Stanford News. Retrieved December 5, 2024, from https://sustainability.stanford.edu/news/understanding-how-sunscreens-damage-coral
Kerlin, K. (2021, March 31). Seagrasses turn back the clock on ocean acidification. University of California Davis. Retrieved November 22, 2024, from https://www.ucdavis.edu/climate/news/seagrasses-turn-back-the-clock-on-ocean-acidification
Kikkert, J. R., Vidal, J. R., & Reisch, B. I. (2005). Stable transformation of plant cells by particle bombardment/biolistics. Methods in molecular biology (Clifton, N.J.), 286, 61–78. https://doi.org/10.1385/1-59259-827-7:061
Levin, R. A., Voolstra, C. R., Agrawal, S., Steinberg, P. D., Suggett, D. J., & van Oppen, M. J. H. (2017). Engineering strategies to decode and enhance the genomes of coral symbionts. Frontiers in microbiology, 8, 1220. https://doi.org/10.3389/fmicb.2017.01220
Lindsey, R., & Dahlman, L. (2024, January 18). Climate change: Global temperature National Oceanic and Atmospheric Administration. Retrieved December 3, 2024, from https://www.climate.gov/news-features/understanding-climate/climate-change-global-temperature#:~:text=According%20to%20the%202017%20U.S.,up%20to%205.9%20degrees%20warmer.
Montalbetti, E., Biscéré, T., Ferrier-Pagès, C., Houlbrèque, F., Orlandi, I., Forcella, M., Galli, P., Vai, M., & Seveso, D. (2021). Manganese benefits heat-stressed corals at the cellular level. Frontiers, 8. https://doi.org/10.3389/fmars.2021.681119
Monterey Bay Aquarium. (n.d.). How seagrass and kelp support habitats’ resilience in a changing ocean. https://www.montereybayaquarium.org/stories/seagrass-kelp-help-climate-change-ocean-acidification
National Geographic Society. (2024, November 21). Coral. https://education.nationalgeographic.org/resource/coral/
National Oceanic and Atmospheric Administration Coral Reef Information System (NOAA CoRIS). (2025, February 28). What are coral reefs. Retrieved November 22, 2024. https://www.coris.noaa.gov/about/what_are/
National Park Service. (2022, October 5). Oceans, coasts & seashores. Retrieved December 3, 2024, from https://www.nps.gov/subjects/oceans/sunscreen.htm
Reef Resilience Network. (n.d.). Mass bleaching. Retrieved November 20, 2024. https://reefresilience.org/bleaching/mass-bleaching/
Shinescreen. (n.d.). IGem. Retrieved November 14, 2024, from https://2021.igem.org/Team:St_Andrews
Swindell, W. R., Huebner, M., & Weber, A. P. (2007). Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC genomics, 8, 125. https://doi.org/10.1186/1471-2164-8-125
Team: GA State SW Jiaotong. (2020). GA State SW Jiaotong iGEM Team. iGEM. Retrieved April 3, 2025, from https://2020.igem.org/Team:GA_State_SW_Jiaotong/Engineering
United Nations Environment Programme. (2025, January 20). Coral reefs. Retrieved November 14, 2024. https://www.unep.org/topics/ocean-seas-and-coasts/blue-ecosystems/coral-reefs
Urry, A. (2015, November 22). Coral reefs are in trouble. Meet the people trying to rebuild them. Vox. Retrieved January 9, 2025, from https://www.vox.com/2015/11/22/9778494/coral-reef
Voolstra, C. R., Aranda, M., Zhan, Y., & Dekker, J. (2021). Symbiodinium microadriaticum (coral microalgal endosymbiont). Trends in Genetics, 37(11). https://doi.org/10.1016/j.tig.2021.08.008
Voolstra, C. R., & Ziegler, M. (2020). Adapting with microbial help: Microbiome flexibility facilitates rapid responses to environmental change. BioEssays : news and reviews in molecular, cellular and developmental biology, 42(7), e2000004. https://doi.org/10.1002/bies.202000004
Wang, L., Ma, K. B., Lu, Z. G., Ren, S. X., Jiang, H. R., Cui, J. W., Chen, G., Teng, N. J., Lam, H. M., & Jin, B. (2020). Differential physiological, transcriptomic and metabolomic responses of Arabidopsis leaves under prolonged warming and heat shock. BMC plant biology, 20(1), 86. https://doi.org/10.1186/s12870-020-2292-y
Withers, A., & Dickie, G. (2024, October 17). Exclusive: Global coral bleaching event expands, now the largest on record. Reuters. Retrieved November 14, 2024, from https://www.reuters.com/business/environment/global-coral-bleaching-event-expands-now-largest-record-2024-10-17/