Ajai Bubra, Emma van Ginkel, and Brenna Pethel, Western Reserve Academy, Hudson, Ohio, United States
Reviewed on 3 May 2025; Accepted on 9 June 2025; Published on 27 October 2025
With help from the 2025 BioTreks Production Team.
Greenhouse gas emissions and food waste are critical global challenges that exacerbate climate change, environmental degradation, and public health risks. Fruit waste, in particular, significantly contributes to greenhouse gas emissions due to methane release during decomposition. Current renewable energy solutions, such as electric vehicles, offer partial relief from CO2 emissions; however, they are hindered by high costs and environmentally damaging lithium mining. This study proposes a synthetic biology approach to convert fruit waste into bioethanol, a cleaner-burning fuel that reduces greenhouse gas emissions by up to 48%. By genetically engineering Escherichia coli with the pyruvate decarboxylase (pdc) and alcohol dehydrogenase (adhB) genes from Zymomonas mobilis, we aim to enhance microbial ethanol production. Z. mobilis is selected for its effective use of the adhb and pdc genes in rapid sugar metabolism. In the past, Z. mobilis was used to produce alcoholic beverages due to its fermentation efficiency. Z. mobilis also produces minimal byproduct formation and high ethanol yield. Because fruit waste contains complex carbohydrates such as cellulose and pectin, enzymatic pretreatment with cellulolytic and pectinolytic enzymes is necessary to release fermentable sugars. The engineered E. coli quickly metabolizes these sugars into ethanol and carbon dioxide. The residual biomass can be repurposed as livestock feed, promoting a circular bioeconomy. This scalable and cost-effective approach transforms agricultural waste into a renewable energy source, mitigating food waste and reducing reliance on fossil fuels. By integrating synthetic biology with waste valorization, this project presents an innovative and sustainable solution to energy production, demonstrating the potential of biotechnology to advance global sustainability efforts.
Keywords: Fruit waste, bioethanol, Zymomonas mobilis, Escherichia coli, pyruvate decarboxylase, alcohol dehydrogenase
Authors are listed in alphabetical order. Gitanshu Bhatia and Beth Pethel mentored the group. Please direct all correspondence to pethelb@wra.net.



Background
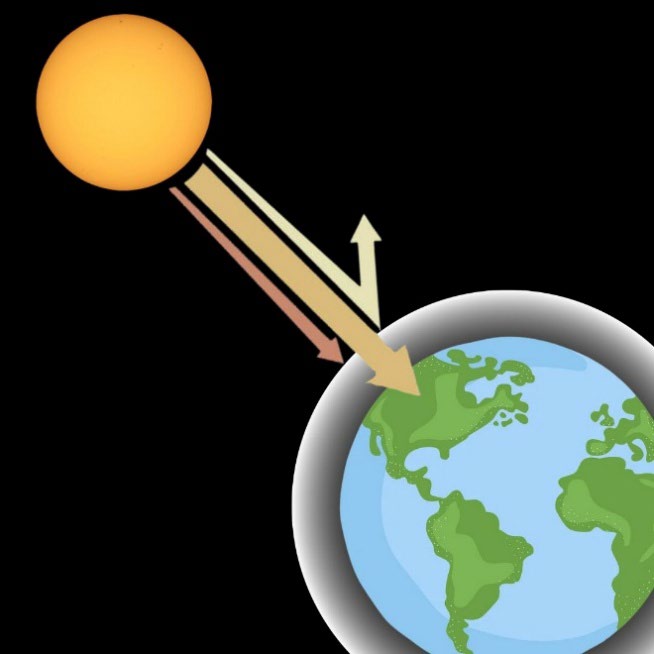
Greenhouse gases such as carbon dioxide (CO₂), methane (CH₄), nitrous oxide (N₂O), chlorofluorocarbons, and hydrofluorocarbons are among the most significant contributors to global warming, posing a large threat to planetary climate stability and human health. Although found in smaller concentrations than CO₂, Methane is over 25 times more effective at trapping heat in the atmosphere over 100 years, making it a particularly potent greenhouse gas. Up to 58% of methane comes from food waste (U.S. Environmental Protection Agency, 2023). These gases trap the sun’s radiation in the Earth’s atmosphere, leading to continually rising global temperatures (Figure 1). As a result, the 2011-2020 period was the hottest decade on record, emphasizing the need to reduce greenhouse gas emissions (Causes and Effects of Climate Change, 2024).
| Figure 1. A diagram illustrating the greenhouse effect shows that greenhouse gases in the Earth’s atmosphere trap heat, preventing it from escaping back into space and contributing to global warming. |
|
The decomposition of fruit waste contributes largely to the amount of methane in the air. Around 20–25% of the world’s yearly fruit production is left out to decompose, creating a critical environmental issue (Radzyminski, 2020). Anaerobically decomposed, the waste releases methane into the environment, increasing the rate at which heat is trapped in the atmosphere.
The consequences of rising global temperatures include increased atmospheric water vapour, resulting in more intense rain, flooding, and intense storms across regions. Global warming affects access to water, disrupts valuable resources like fisheries and agriculture, and worsens global food insecurity. Since 1910, the United States alone has experienced an increase in precipitation by .5%each decade (U.S. Environmental Protection Agency, 2024). Besides inflicting harm on the environment, global warming remains a major public health threat, increasing air pollution rates, disease transmission, mental stress, and the rates of hunger and malnutrition. Globally, these negative environmental factors result in about 13 million deaths yearly.
In the United States, almost 80% of transportation-related greenhouse gas emissions stem from the combustion of gasoline and diesel (Causes and Effects of Climate Change, 2024). In an attempt to mitigate these emissions, engineers created several technologies like electric vehicles, carbon capture systems, and clean energy. However, electric cars exist only through the mining of lithium-ion batteries, the process of which pollutes the water and air, and carbon capture remains energy and cost-intensive (Biello, 2010; Stromberg, 2013). Addressing these issues will lead to sustainable energy options for mass populations.
With the help of advances in synthetic biology, our project aims to utilize fruit waste, an untapped bioresource, by converting organic fruit waste into ethanol biofuel. Not only does this address food waste, but it also becomes a cheap renewable energy source. Because the family of one of our team members (AB) runs a Blueberry farm and witnesses the wasted potential firsthand, we decided to confront the issue of greenhouse gas emissions by taking advantage of fruit waste..
To create a cost-effective and clean energy source, we will genetically engineer Escherichia coli by adding pyruvate decarboxylase (pdc) and alcohol dehydrogenase (adhB) genes from Zymomonas mobilis, a bacterium notable for its high ethanol production efficiency (Ingram, 1987). Although Z. mobilis possesses a greater ethanol production than that of the majority of microorganisms, it lacks the metabolic stability and versatility to ferment complex biomass hydrolysates. E.coli is also preferred due to its promoter strength and industrial characteristics, allowing it to be a better chassis for our design. E. coli can break down a broader range of sugars, such as glucose, fructose, arabinose, and xylose, allowing for a more effective fermentation process, counteracting Z. mobilis’s deficiencies and making it a more convenient industrial microorganism to utilize when generating biofuel on a large scale (Turner, 2012; “Zymomonas mobilis,” 2014). With our design, we aim to further enhance the fermentation and bioethanol production of Z. mobilis by using pdc and adhB, genes in Z. mobilis which are known to be the backbone for Z. mobilis’ bioethanol production (U. Kalnenieks, 2006). Our project aims to optimize bioethanol production by placing pdc and adhB under a dual promoter system that responds in the presence of fructose and glucose.
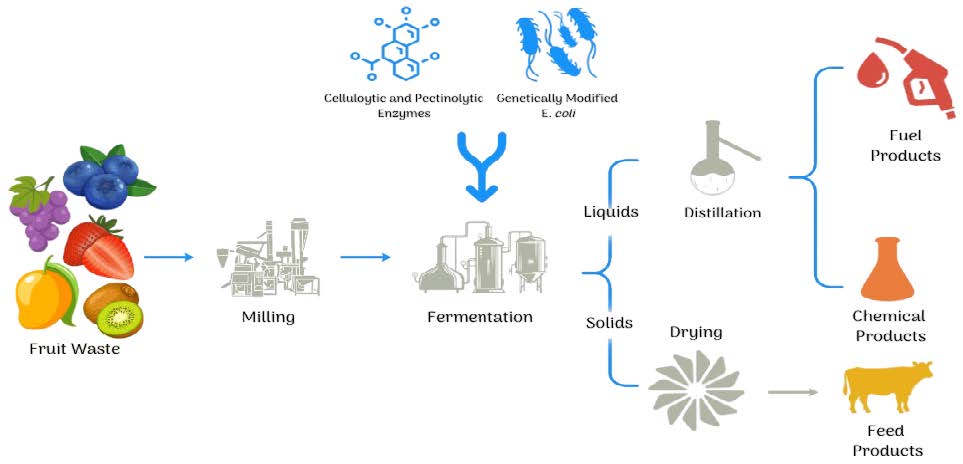
Our process begins by obtaining fruit waste from the local farms, milling, and pretreating the waste with cellulolytic and pectinolytic enzymes under anaerobic conditions. The anaerobic conditions will help the enzymes break down cellulose and pectin, two prominent structural carbohydrates in plant cell walls, into fermentable sugars such as glucose (JGI Genome Portal, n.d.; Tungeker, 2021). The engineered E. coli then ferments the obtained glucose to produce ethanol and carbon dioxide. As evident in Figure 2, the produced ethanol can be filtered and used as an alternative clean-burning fuel (Stromberg, 2013; Biello, 2010). The remaining plant material after fermentation can be dried and used as animal feed, creating a sustainable, recyclable cycle (Shehu, 2019). This dual-purpose approach provides a single solution to two pressing concerns: methane emissions from organic waste and the global need for cleaner energy resources.
| Figure 2. Illustrates a holistic view of the process of turning fruit waste into biofuel. Starting on the left, the fruit waste goes through an initial milling phase in which it is crushed and mixed, turning it to almost a jam-like consistency to release juices. Next, the milled fruit waste enters an anaerobic vat and is exposed to cellulolytic and pectinolytic enzymes to break down pectin and cellulose. Once the cellulose and pectin are broken down, we add in our engineered E. coli which will ferment and efficiently produce bioethanol. Once the E. coli has fermented the fruit waste, the liquids and solids are separated outside of the anaerobic chamber, where the liquids are distilled into bioethanol and used for vehicles while the remaining fruit waste solids, skins and seeds, are dried and re-fed to livestock. |
|
Synthetic biology thereby provides an efficient solution for pressing environmental issues.
Because of advanced metabolic engineering technology, microbial ethanol production has become highly efficient, making E. coli, with its genetic adaptability and programmability, a lead organism in the biofuel industry (Dien, 2003). By reducing climate change and food waste, and producing renewable energy, microbial ethanol production presents a promising, clean solution to the global overproduction of greenhouse gases (Causes and Effects of Climate Change, 2024).
Systems level
Designed to address greenhouse gas emissions and food waste, our project turns fruit waste into a clean-burning biofuel by converting fruit biomass into ethanol using genetically engineered Escherichia coli. It involves the enzymatic treatment of fruit waste, anaerobic microbial fermentation, the recovery of ethanol, and biomass recycling (Saratale, 2018; Singh, 2017). Fruit waste is rich in complex carbohydrates such as cellulose and pectin, which turn into fermentable sugars, such as glucose, with the aid of cellulolytic and pectinolytic enzymes (Kaur, 2020). Engineered E. coli utilizes those sugars and produces ethanol through an adjusted glycolytic pathway with pyruvate decarboxylase (pdc) and alcohol dehydrogenase (adhB), genes sourced from the bacterium Zymomonas mobilis (Ingram, 1987; Gunasekaran and Raj, 2001). The resulting ethanol is recovered, and the residual biomass can be recycled as feed for livestock (Bhatia, 2022).
Device level
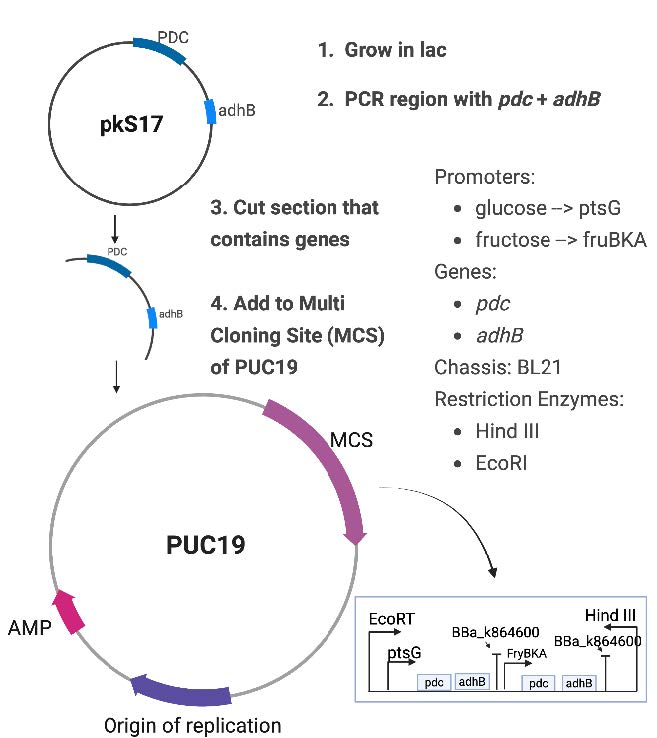
Our microbial system utilizes a dual-promoter synthetic plasmid design to promote bioethanol yield (Green & Sambrook, 2012). To start, plasmid pKS17, which consists of both pdc and adhB genes, is used to amplify both genes into a single gene fragment (Figure 3). Following PCR amplification, the gene fragment containing both pdc and adhB genes is cut with the restriction enzymes EcoRI and HindIII and cloned into the plasmid pUC19 MCS.
| Figure 3. A visual representation of our two-plasmid process to express pdc and adhB into E. coli, with a closer look at our promoters and restriction enzymes. One promoter is glucose-induced while the other is fructose-induced, allowing for optimal fermentation. The legend illustrates the genes, chassis, promoters, and restriction enzymes that we are using. |
|
The constructed pUC19 construct utilizes EcoRI and HindIII sites for directional cloning. A glucose-inducible ptsG promoter precedes the pdc and adhB genes in the construct, followed by the terminator BBa_K864600. A fructose-inducible fruBKA promoter controls a second pdc and adhB expression unit terminated by BBa_K864600. The two-promoter system allows for carbon source-regulated ethanol production (Figure 4).
The pUC19 plasmid, used for its efficiency and versatility in genetic engineering, is transformed into E. coli BL21, a strain engineered for high levels of gene and protein expression.
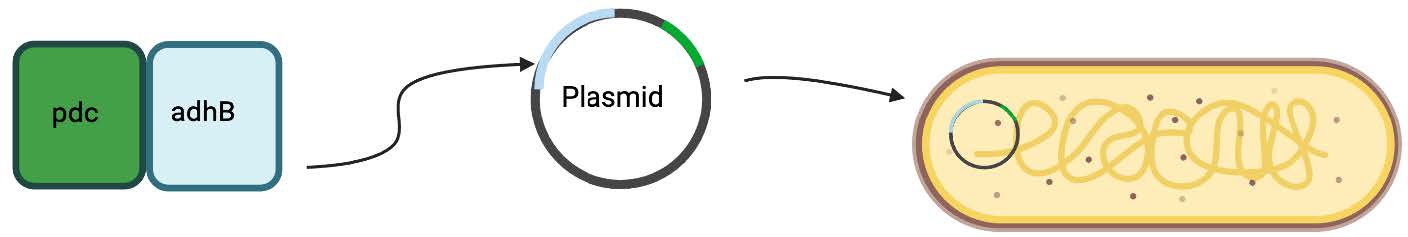
| Figure 4. This figure represents the process of extracting the pdc and adhB genes, using our two-plasmid and two-promoter method, and inserting the plasmid into our chassis. |
|
This strain will be cultured anaerobically with enzymatically treated fruit waste, ensuring efficient ethanol production from both the glucose and fructose substrates present.
Parts level
The process integrates important molecular components for accurate gene control and high-yield ethanol production. The pdc gene encodes pyruvate decarboxylase, which converts pyruvate to acetaldehyde and CO₂, and adhB encodes alcohol dehydrogenase (Ingram et al., 1987), which converts acetaldehyde to ethanol (Ingram, 1987) (Figure 5).
| Figure 5. Chemical production of ethanol: As pyruvate is a product of glycolysis, the enzyme pyruvate decarboxylase allows for a chemical conversion into acetaldehyde (Candy, J. M., 1998). The acetaldehyde is an intermediate molecule that, when exposed to alcohol dehydrogenase, can be converted to our final product of ethanol. |
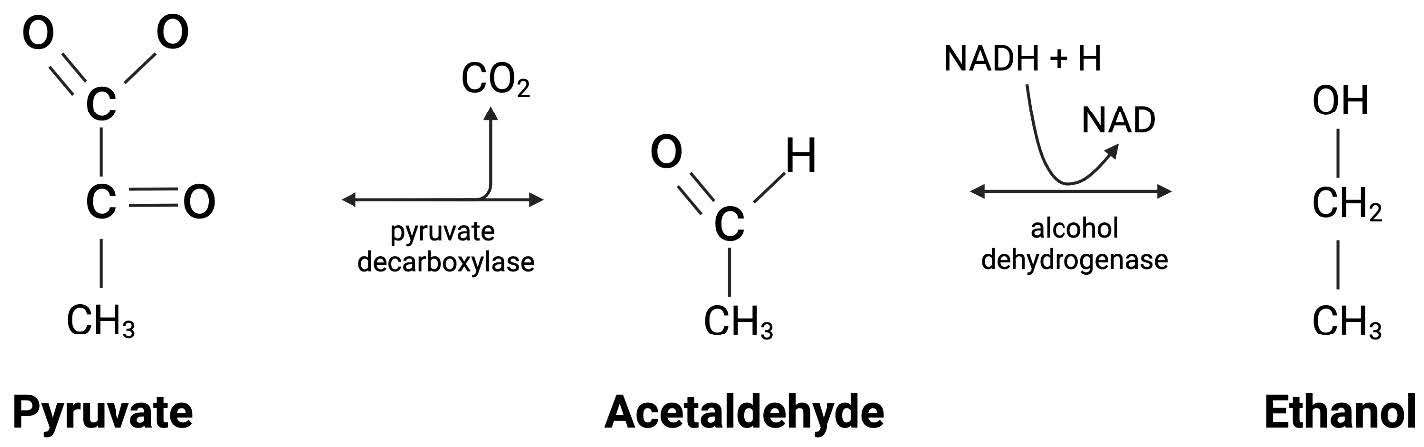
|
Two carbon source-regulated promoters control these genes: fruBKA for fructose and ptsG for glucose. The two transcriptional units contain a strong bacterial terminator (BBa_K864600) for efficient transcription termination. Cloning is achieved by using EcoRI (NEB #R0101) and HindIII (NEB #R0104), which generate compatible 5′ overhangs. T4 DNA ligase allows for ligation, and the final plasmid construct containing the two transcriptional units is transformed into E. coli BL21 cells. Antibiotic resistance results in success, and PCR and DNA sequencing confirm gene insertion.
Safety
Although the fermentation process in our design produces an ethanol concentration sufficient to kill E. coli (in the 60–90% alcohol range), release of genetically modified bacteria into the biosphere remains a significant health and safety concern (Madigan, 2015). Ethanol breaks down the bacterial cell membrane, effectively lysing the E. coli during fermentation (Ingram, 1987). However, there remains a possibility that the bacteria escapes into the environment before completely lysis, incidentally transferring drug-resistant plasmids to native microorganisms through horizontal gene transfer (Davison, 1999; Thomas & Nielsen, 2005). Using tetracycline as a selection marker, which makes E. coli resistant to antibiotics, heightens this risk (Chopra and Roberts, 2001). Adherence to rigid containment of bacteria within the fermentor system and enforcement of proper personal protective equipment (PPE) will reduce the potential for inadvertent release. Sterilization using chemicals for all biological wastes and prohibition against discharging wastes to municipal sewage systems will minimize environmental release of genetically modified organisms (GMOs) and antibiotic-resistant genes (EPA, 2012). Final bioethanol will undergo testing to ensure there is no contamination of E.coli.
Pdc and adhB, the enzymes engineered into E. coli to aid in fruit biomass breakdown, can act as allergens or irritants if released into the air (Kim, 2006). To mitigate this, we will handle all enzymes with proper personal protective equipment and work in fume hoods when necessary.
Over time, engineered E. coli can lose plasmids or become mutated, reducing ethanol yield or generating undesired byproducts (Silva-Rocha and de Lorenzo, 2010). Regularly monitoring fermentation products, regular plasmid stability tests, and sequencing verification will maintain system integrity.
Ethanol’s high flammability poses serious safety hazards, so following fire safety guidelines by using explosion-proof containers and fume hoods and monitoring ethanol concentrations will keep the fermentation process safe and efficient.
Fruit waste’s composition may vary, affecting fermentation consistency and possibly generating unknown byproducts such as mold and pathogenic bacteria. To avoid this, the fruit waste will be prescreened and regulated.
By carefully testing our design, implementing thorough containment strategies, and following safety guidelines, we can ensure that this project will not harm people or the environment.
Discussions
Integrating synthetic biology into biofuel production presents a promising solution to the dual environmental challenges of greenhouse gas emissions and food waste. Our project demonstrates a sustainable approach by utilizing genetically engineered Escherichia coli to convert enzymatically treated fruit biomass into ethanol. This method gives organic waste value and mitigates methane emissions typically associated with fruit decomposition.
The use of Zymomonas mobilis-derived pdc and adhb genes leverages well-characterized enzymatic efficiency in ethanol biosynthesis (Boismier, E. C., 2025). These genes, incorporated into E. coli BL21, enable the redirection of metabolic flux from pyruvate toward ethanol through a simplified and efficient pathway. We selected BL21 for its high-level protein expression capabilities, which maximize ethanol output under anaerobic conditions.
A key innovation in our system lies in the dual-promoter plasmid design. By embedding the pdc and adhB genes under glucose-inducible (ptsG) and fructose-inducible (fruBKA) promoters, our design allows for carbon source-responsive expression. This regulatory construct aligns with the carbohydrate composition of fruit hydrolysates, which are rich in glucose and fructose. The dual-promoter system optimizes ethanol production by activating gene expression based on available substrates, enhancing the metabolic flexibility of the chassis organism.
Moreover, the modular assembly strategy offers a versatile cloning framework, beginning with gene amplification in plasmid pKS17 and subsequent integration into pUC19. Using applicable restriction sites (EcoRI and HindIII) and strong terminators (BBa_K864600) ensures precise gene insertion and controlled transcription termination, critical for reliable expression.
The reuse of fermentation residues as livestock feed supports a circular bioeconomy. This reduces environmental waste but also provides secondary economic value, contributing to the system’s scalability and sustainability.
Despite the advantages, several challenges remain. Efficient hydrolysis of complex carbohydrates requires cost-effective enzyme formulations, and large-scale fermentation may introduce variability in substrate composition and microbial performance. Future work should include optimization of fermentation conditions, long-term plasmid stability assessments, and quantitative evaluation of ethanol yield across different fruit waste inputs.
Overall, this project exemplifies the potential of synthetic biology in sustainable energy innovation. By integrating waste management with renewable fuel production, we advance toward a more circular and resilient bioeconomy.
Next steps
To test and execute our design, we will start by collecting fruit waste from farmers, restaurants, grocery stores, et cetera and crushing all the waste together to treat it enzymatically using cellulolytic and pectinolytic enzymes under anaerobic conditions, releasing glucose and fructose–fermentable sugars. We will then complete the process of engineering a dual-promotor plasmid, highlighted in the Device level and Parts level sections, by inserting Zymomonas mobilis genes into E. coli BL21. These genes will be placed under the control of glucose and fructose-inducible promoters to enable ethanol production. PCR amplification followed by the transfer of plasmid pUC19 into the MCS and EcoRI and HindIII sites, joining the engineered pUC19, will enable cloning. The engineered bacteria will be cultured anaerobically with pretreated fruit mass, consequently converting sugars into ethanol. Ethanol will then be extracted, and the remaining biomass will be tested for live E. coli before being repurposed as livestock feed to promote sustainability. We will follow all safety guidelines found in the Safety section to create a sustainable fuel alternative.
Author contributions
AB thought of the concept, designed the study, completed in-depth research, and wrote the Discussions section. BP created images and the video, and kept track of references. EV created images, wrote Next Steps and Safety, and thoroughly reviewed all written works. All worked collaboratively to write the Systems, Device, and Parts level sections, as well as the Background and Abstract.
Acknowledgments
We would like to thank Dr. Pethel for providing valuable guidance, feedback, and expertise while creating this project. We extend our gratitude to Gitanshu Bhatia, our BioBuilder mentor, for thoughtfully reviewing our work and offering constructive comments. Finally, we acknowledge BioTreks for their support in making this study possible.
References
Addgene. (n.d.). Plasmid #24628: pKS17. Retrieved March 2025, from https://www.addgene.org/24628
Bhatia, S. K., et al. (2022). Bioresource Technology Reports, 19, 101171. https://doi.org/10.1016/j.biteb.2022.101171
Biello, D. (2010). Are electric cars really so green? Scientific American. Retrieved from https://www.scientificamerican.com
Biello, D. (2010, January 27). Bacteria transformed into biofuel refineries. Scientific American. Retrieved November 19, 2024, from https://www.scientificamerican.com/article/bacteria-transformed-into-biofuel-refineries/
Boismier, E. C., Aboulnaga, E. A., & TerAvest, M. A. (2025). Zymomonas mobilis: Bringing an ancient human tool into the genomic era. Current Opinion in Biotechnology, 92, 103257. https://doi.org/10.1016/j.copbio.2025.103257
Candy, J. M. (1998, December 23). Structure and properties of pyruvate decarboxylase and site-directed mutagenesis of the Zymomonas mobilis enzyme. ScienceDirect. Retrieved November 19, 2024, from https://www.sciencedirect.com/science/article/abs/pii/S0167483898000776?via%3Dihub
Chopra, I., & Roberts, M. (2001). Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiology and Molecular Biology Reviews, 65(2), 232–260. https://doi.org/10.1128/MMBR.65.2.232-260.2001
Davison, J. (1999). Genetic exchange between bacteria in the environment. Plasmid, 42(2), 73–91. https://doi.org/10.1006/plas.1999.1421
Dien, B. S. (2003, December 16). Bacteria engineered for fuel ethanol production: Current status. Springer Nature Link. Retrieved November 19, 2024, from https://link.springer.com/article/10.1007/s00253-003-1444-y
Dien, B. S., Cotta, M. A., & Jeffries, T. W. (2003). Bacteria engineered for fuel ethanol production: Current status. Applied Microbiology and Biotechnology, 63, 258–266. https://doi.org/10.1007/s00253-003-1444-y
Environmental Protection Agency. (2012). Guidance for containment and control of genetically modified microorganisms (EPA Publication No. EPA-600-R-12-019). U.S. Environmental Protection Agency. https://nepis.epa.gov/Exe/ZyPDF.cgi/P100F4ZI.PDF
EPA. (2012). Guidance for containment and disposal of genetically engineered microorganisms. https://www.epa.gov/sites/default/files/2015-06/documents/biotech_guidance.pdf
Frequently Asked Questions (FAQs) – U.S. Energy Information Administration. (n.d.). U.S. EIA. Retrieved November 19, 2024, from https://www.eia.gov/tools/faqs/faq.php?id=307&t=10
Green, M. R., & Sambrook, J. (2012). Molecular cloning: A laboratory manual. Cold Spring Harbor Laboratory Press. https://www.cshlpress.com/default.tpl?cart=17132371093573390&fromlink=T&linkaction=full&linksortby=oop_title&–eqSKUdatarq=9781936113422
Gunasekaran, P., & Raj, K. C. (2001). Journal of Biosciences, 26(4), 415–426. https://doi.org/10.1007/BF02704642
Ingram, L. O. (1987). Changes in lipid composition of Escherichia coli resulting from growth with ethanol. Journal of Bacteriology, 151(2), 891–897. https://doi.org/10.1128/jb.151.2.891-897.1987
Ingram L. O., Conway T., Clark D. P., Sewell G. W., Preston J. F. Genetic engineering of ethanol production in Escherichia coli. Appl Environ Microbiol. 1987 Oct;53(10):2420-5. doi: 10.1128/aem.53.10.2420-2425.1987. PMID: 3322191; PMCID: PMC204123.
Ingram, L. O., Aldrich, H. C., Borges, A. C., Causey, T. B., Martinez, A., Morales, F., Saleh, A., Underwood, S. A., Yomano, L. P., York, S. W., Zaldivar, J., & Zhou, S. (1999). Enteric bacterial catalysts for fuel ethanol production. Biotechnology Progress, 15(5), 855–866. https://doi.org/10.1021/bp990110w
Ingram, L. O., et al. (1987). Ethanol production by recombinant Escherichia coli strains coexpressing Zymomonas mobilis genes encoding pyruvate decarboxylase and alcohol dehydrogenase. Applied and Environmental Microbiology, 53(10), 2420–2425. https://aem.asm.org/content/53/10/2420
JGI Genome Portal. (n.d.). Pectinase and cellulase gene resources. Retrieved from https://genome.jgi.doe.gov
Kaur, H., et al. (2020). Waste and Biomass Valorization, 11, 5335–5350. https://doi.org/10.1007/s12649-019-00773-0
Kim, J. H., et al. (2006). Occupational allergy due to enzymes in biotechnology and food industries. Annals of Allergy, Asthma & Immunology, 97(1), 108–112. https://doi.org/10.1016/S1081-1206(10)61379-1
Madigan, M. T., et al. (2015). Brock biology of microorganisms. Pearson. https://www.pearson.com/store/p/brock-biology-of-microorganisms/P100000219183
New England Biolabs. (n.d.). EcoRI-HF® (R3101) and HindIII-HF® (R3104) Product Information. Retrieved March 2025, from https://www.neb.com
PLOS Blogs. (2016, May 16). iGEM REPORT: From carbon dioxide to biofuel. Retrieved November 19, 2024, from https://collectionsblog.plos.org/igem-report-17-01/
Radzyminski, K. (2020). The impact of food waste on climate change. Journal of Sustainability and Environmental Management, 8(3), 112–119.
Sambrook, J., & Russell, D. W. (2001). Molecular cloning: A laboratory manual. Cold Spring Harbor Laboratory Press. https://www.cshlpress.com/default.tpl?cart=17132371093573390&fromlink=T&linkaction=full&linksortby=oop_title&–eqSKUdatarq=587
Saratale, R. G., et al. (2018). Renewable and Sustainable Energy Reviews, 82, 3311–3326. https://doi.org/10.1016/j.rser.2017.10.041
Shehu, I. (2019, October 14). Fruit, nut, cereal, and vegetable waste valorization to produce biofuel. Wiley. Retrieved November 19, 2024, from https://onlinelibrary.wiley.com/doi/10.1002/9781119383956.ch30
Silva-Rocha, R., & de Lorenzo, V. (2010). Noise and robustness in prokaryotic regulatory networks. Annual Review of Microbiology, 64, 257–275. https://doi.org/10.1146/annurev.micro.112408.134306
Singh, R., et al. (2017). Bioresource Technology, 245, 1120–1129. https://doi.org/10.1016/j.biortech.2017.08.213
Strategies to Optimize Protein Expression in E. coli. (2010, August 1). Current Protocols. Retrieved November 19, 2024, from https://currentprotocols.onlinelibrary.wiley.com/doi/10.1002/0471140864.ps0524s61
Stromberg, J. (2013). Genetically modified E. coli bacteria can now synthesize diesel fuel. Smithsonian Magazine. Retrieved November 19, 2024, from https://www.smithsonianmag.com/science-nature/genetically-modified-e-coli-bacteria-can-now-synthesize-diesel-fuel-38362260/
Stromberg, J. (2013, April 22). The limits of carbon capture. Smithsonian Magazine. Retrieved November 19, 2024, from https://www.smithsonianmag.com
Thomas, C. M., & Nielsen, K. M. (2005). Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nature Reviews Microbiology, 3(9), 711–721. https://doi.org/10.1038/nrmicro1234
Turner, P. C. (2012, April 1). Optical mapping and sequencing of the Escherichia coli KO11 genome. Oxford Academic. Retrieved November 19, 2024, from https://academic.oup.com/jimb/article/39/4/629/5994587
Tungekar, S. D. (2021). Cellulolytic enzymes and microbial conversion of biomass to bioethanol. BioResources, 16(2), 2849–2866.
U. Kalnenieks, N. Galinina, M.M. Toma, J.L. Pickford, R. Rutkis, R.K. Poole. Respiratory behaviour of a Zymomonas mobilis adhB::kanr mutant supports the hypothesis of two alcohol dehydrogenase isoenzymes catalysing opposite reactions, FEBS Letters, Volume 580, Issue 21, 2006, Pages 5084-5088, ISSN 0014-5793, https://doi.org/10.1016/j.febslet.2006.08.034.
U.S. Energy Information Administration. (n.d.). Greenhouse gases and climate change. Retrieved from https://www.eia.gov
U.S. Environmental Protection Agency. (2023, May 25). Quantifying methane emissions from landfilled food waste. https://www.epa.gov/land-research/quantifying-methane-emissions-landfilled-food-waste
U.S. Environmental Protection Agency. (2024, June). Climate change indicators: Heavy precipitation. https://www.epa.gov/climate-indicators/climate-change-indicators-heavy-precipitation
United Nations. (n.d.). Causes and effects of climate change. United Nations. Retrieved from https://www.un.org/en/climatechange/science/causes-effects-climate-change
Wang, D., Gao, Q., & Gao, N. (2015, March 27). Metabolic engineering of Saccharomyces cerevisiae. Annals of Microbiology. https://doi.org/10.1007/s13213-015-1074-5
Wisconsin Team. (2008). Team: Wisconsin/Project. iGem 2008. Retrieved November 19, 2024, from https://2008.igem.org/Team:Wisconsin/Project
Zymomonas mobilis: a novel platform for future biorefineries. (2014, July 2). Biotechnology for Biofuels and Bioproducts. Retrieved November 19, 2024, from http://biotechnologyforbiofuels.biomedcentral.com/articles/10.1186/1754-6834-7-101