Zhongchen Cai, Chun Chan, Ruiwen Wang, and Xinyu You, Western Reserve Academy, Hudson, Ohio, United States
Reviewed on 3 May 2025; Accepted on 9 June 2025; Published on 27 October 2025
With help from the 2025 BioTreks Production Team.
Injuries pose a significant safety issue in daily life. Annually, unintentional injuries cause approximately 200,000 deaths in the United States and affect around 11 million people worldwide. The risks of open wounds arise from bacterial infection and excessive bleeding, causing inflammatory reactions and dilution of healing factors. Current debridement methods that remove damaged tissues from the wound site are unsuitable for large or severely infected wounds, and the lack of healing factors prolongs the wound exposure. Fibrin and fibronectin, naturally found in the human body, are critical for wound healing. Fibrin supports clot formation and cell migration, while fibronectin stimulates angiogenesis, enhances the integrity of the extracellular matrix (ECM), and recruits macrophages to clean the wound site. Unlike fibronectin, which is directly encoded by a gene, fibrinogen must be cleaved by thrombin to form fibrin monomers, which then polymerize. Our project aims to establish a system that simultaneously produces fibronectin, fibrin, and prothrombin, proteins essential for wound healing. The BL21 (DE3) strain of Escherichia coli (E. coli) serves as the chassis for a genetically engineered plasmid that contains genes encoding fibronectin, fibrinogen, and prothrombin. This project will enhance advanced wound care by enabling the stable production of key wound-healing proteins and advancing fibrin cleavage and polymerization, thereby significantly lowering mortality rates and setting a new standard in medical innovation.
Keywords: Wound healing, fibrinogen, fibronectin, fibrin, thrombin, prothrombin, protein production
Authors are listed in alphabetical order. Beth Pethel and Joe Switzer mentored the group. Please direct all correspondence to pethelb@wra.net.


Background
Significance of Wounds
Injuries are a common occurrence in daily life and represent a significant safety concern. Approximately 200,000 people in the United States (U.S.) die annually from unintentional injuries, including burns, falls, and motor vehicle accidents, which rank among the top three causes of death (National Center for Health Statistics [NCHS], 2024). Globally, approximately 11 million people suffer from burn wounds each year (Figures 1 and 2; Our World in Data, 2023; Markiewicz-Gospodarek et al., 2022). Moreover, visits to emergency departments and physicians’ offices for injuries total around 50 million annually (National Center for Health Statistics [NCHS], 2021). In 2019, the estimated economic impact of injuries in the US reached $4.2 trillion (NCHS, 2021).
| Figure 1. Global Distribution of Death Numbers From Fires and Burns in 2021. |
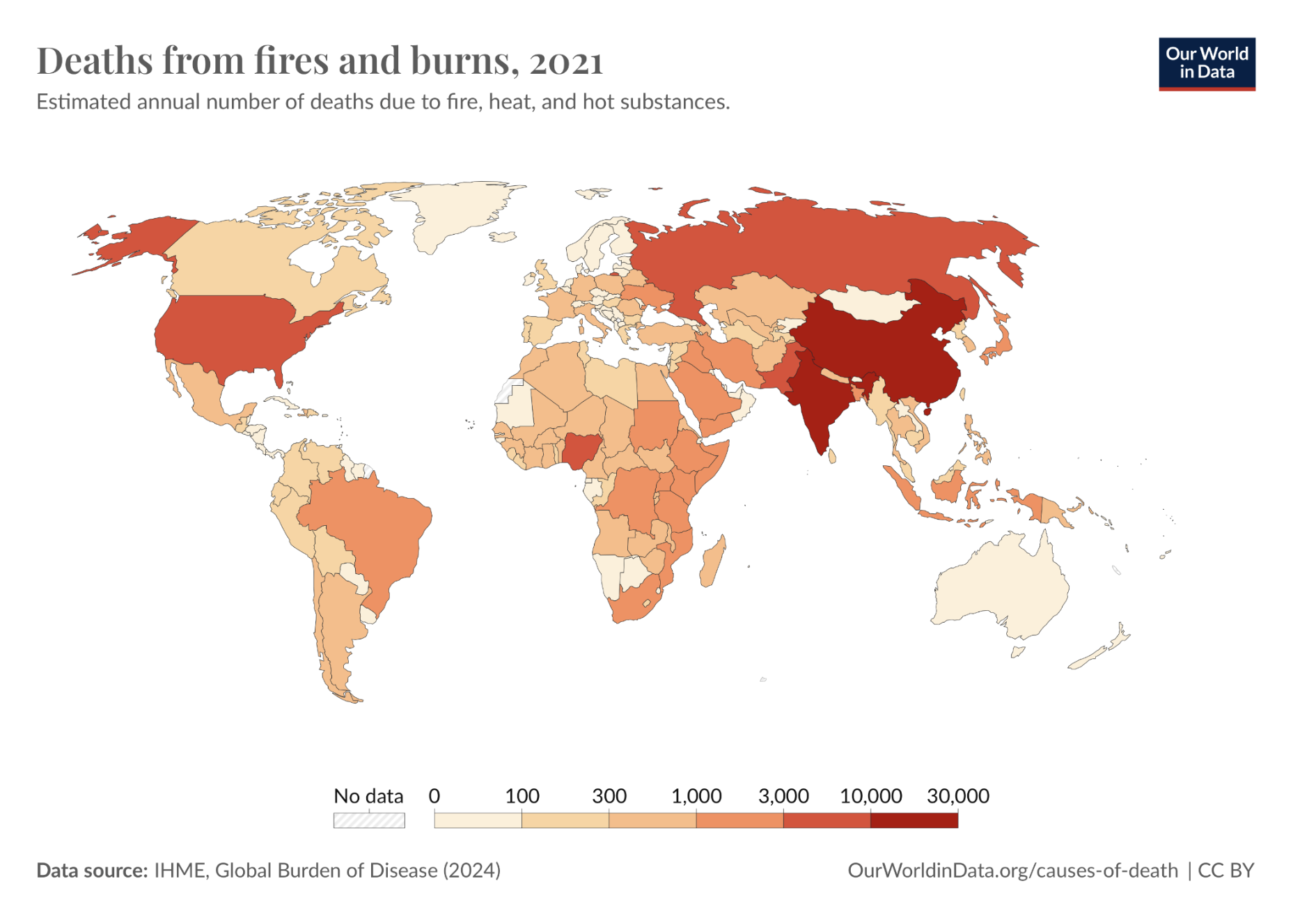
|
This figure illustrates the estimated annual number of deaths resulting from burns. The bar shows the corresponding number represented by the different shading colors, with the higher number displayed in darker red and the lower number displayed in lighter white. The death toll is higher in areas with dense populations, including China, India, and the USA.
World Health Organization (WHO) (2024), with major processing by Our World in Data. “Deaths from fires and burns” [dataset]. World Health Organization, “Global Health Estimates” [original data]. Retrieved May 5, 2025, from https://ourworldindata.org/grapher/deaths-from-fires-and-burns-who
| Figure 2. Death From Fires and Burns. |
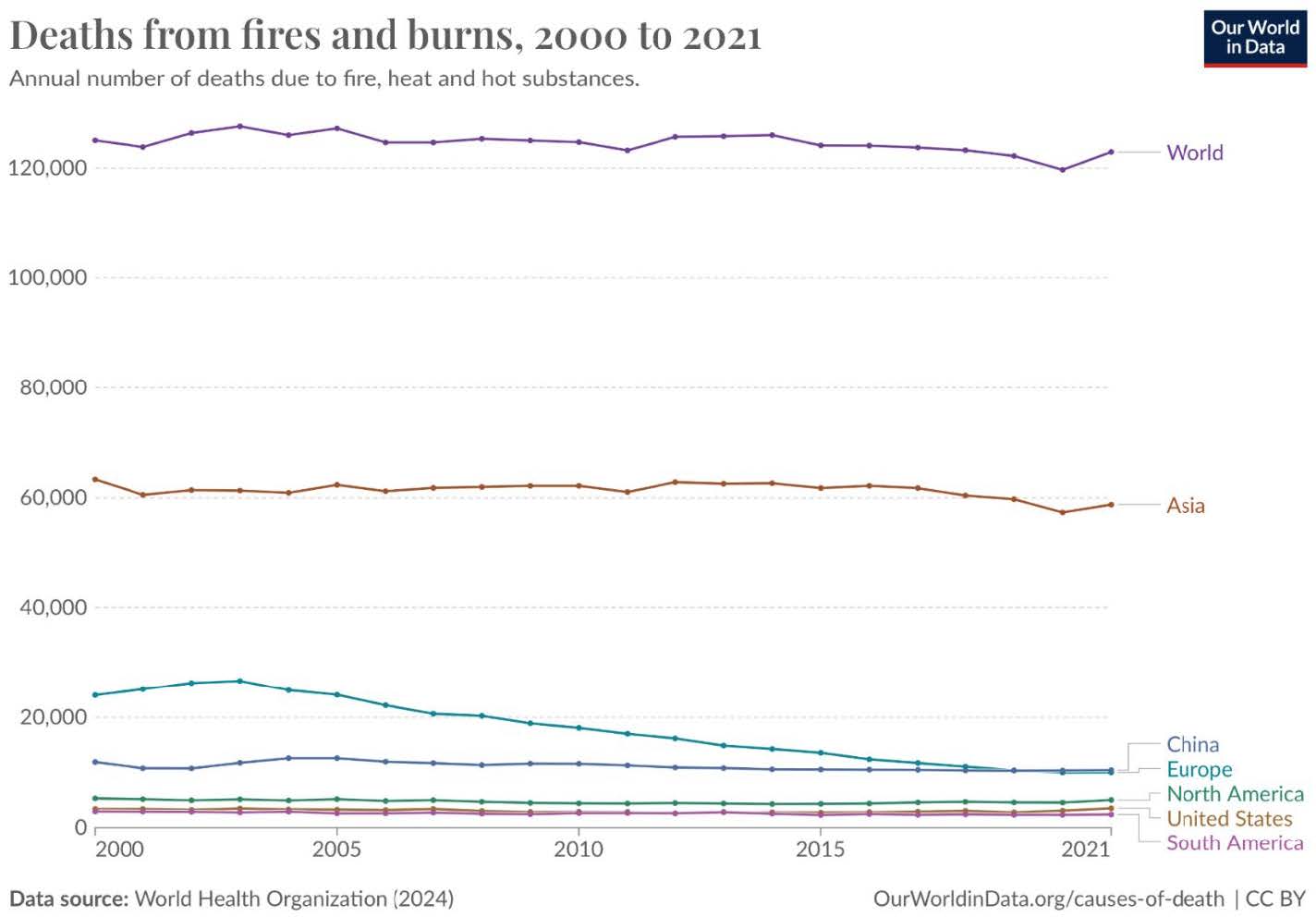
|
Burns and other common injuries pose a significant danger due to their frequent encounters with humans. For instance, daily cookery, flammable chemicals, overexposure to the sun, and combustible materials all lead to potential burning injuries.
Among injuries, blood loss is a leading cause of death in trauma cases (Cleveland Clinic, 2024). Additionally, data from the Trauma Infectious Disease Outcomes Study revealed that over 30 percent of trauma patients experience infections during their initial hospital stay (Tribble et al., 2018).
It takes about two weeks for a healthy person to heal an acute wound, a wound with the proper amount of ECM and Collagen, and that heals at an expected rate. It takes 3 to 4 weeks or longer to heal chronic wounds (wounds with excessive degradation of ECM and Collagen)(Figure 3). Currently, the standard treatments for wound healing include infection treatment, wound debridement, and wound dressing (Fallah et al., 2021).
| Figure 3. Timeline of the Stages of Normal Wound Healing. It takes hours from the hemostasis process to the onset of inflammation. The inflammation process takes days to proceed. The proliferation and remodeling process takes weeks to months. |
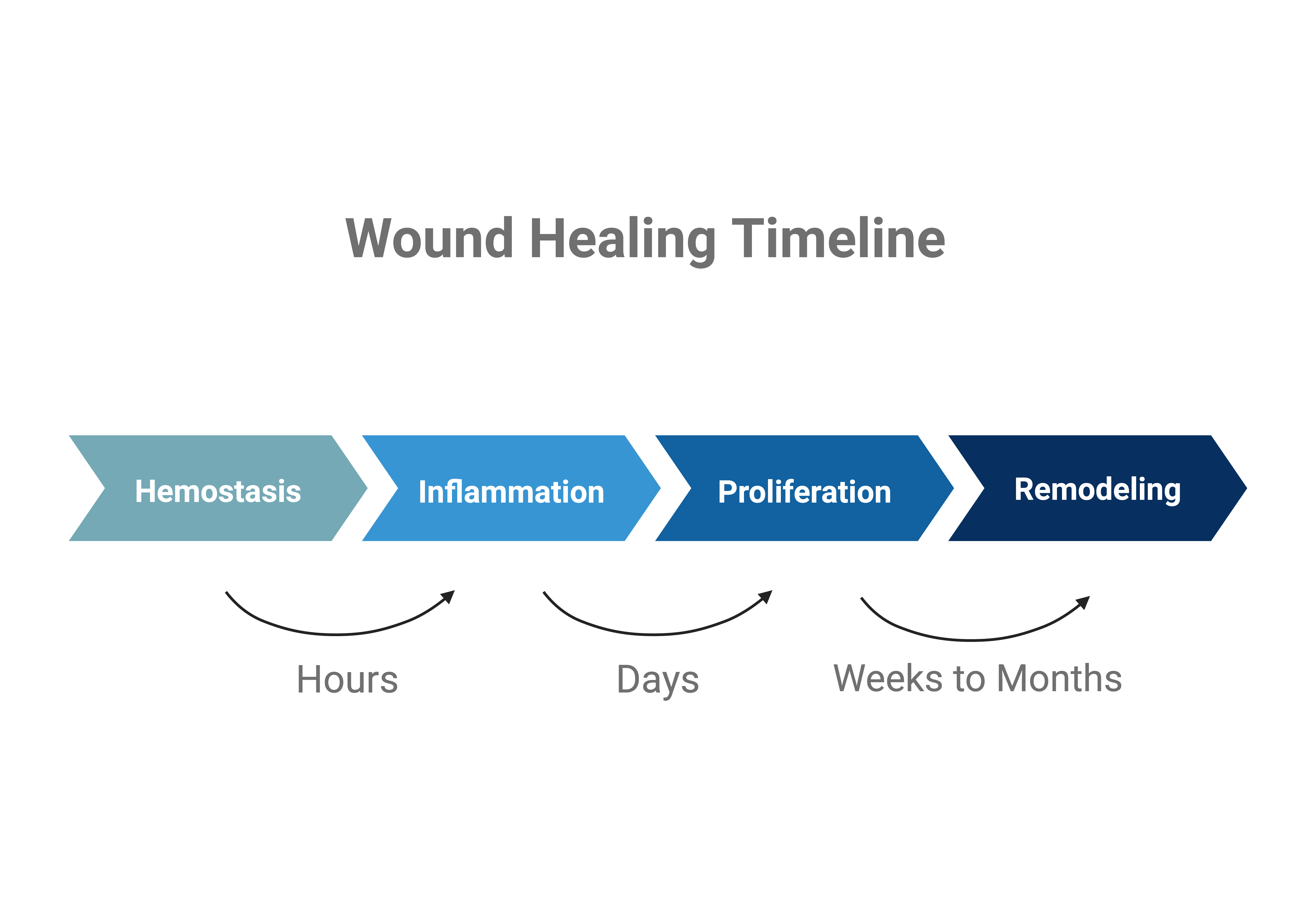
|
Current Wound Treatment Methods
During the wound healing process, several factors delay and pose potential dangers to the wound site. An increased bacterial load around the wound site can disrupt wound healing and lead to infection. Treating bacterial infection is a crucial process during wound healing. Topical cleansers, such as water and saline, as well as antimicrobials like vinegar and acetic acid, neutralize bacteria to prevent and treat bacterial threats to wound healing. Topical antimicrobials are optimal for treating superficial wounds due to their direct contact with bacterial loads, but may lead to bacterial resistance to the antimicrobial agent, resulting in diminished effectiveness as a treatment (Cleveland Clinic, 2024; Drugs.com, 2024; Fallah et al., 2021).
The process of removing damaged tissue, known as wound debridement, plays crucial role in certain cases of wound healing. The presence of dead tissue can interrupt therapy by hindering new tissue growth and blocking keratinocytes (the cells that produce keratin, the main protein in the epidermis) from migrating to the wound bed (Cleveland Clinic, 2024; Fallah et al., 2021). Surgical, mechanical, and enzymatic debridement methods are widely used, employing specific surgical instruments, mechanical forces, and enzymes, respectively, to remove damaged tissues (Fallah et al., 2021). These approaches are unsuitable for large and severely infected wounds (Nunez, 2019). A quick initiation and the speed of the wound healing process reduce the risk of infection, providing the optimal environment for these treatments to address injuries (Kolimi et al., 2022).
Wound dressing refers to the application of materials to promote wound healing. The material types range from bandages to complex engineered stem cells. There are five types of dressings: hydrocolloid, hydrogel, film, alginate, and foam. They are responsible for various wound conditions, including those with different moisture levels and both dry and wet scenarios (Fallah et al., 2021). Each type of dressing is specifically designed for special wound conditions. The addition of fibronectin and fibrin enhances the efficiency of each dressing, amplifying their advantage for a broader range of injury treatment (Kolimi et al., 2022; Lenselink, 2013; Levy et al., 2012; Rousselle et al., 2019; Tan & Dosan, 2019).
The Role of Fibronectin and Fibrin in Wound Healing
The ECM is a network of proteins and other molecules that support and give structure to cells and tissues (National Cancer Institute, n.d.). Fibronectin is the second most abundant ECM molecule in the body (Sicari et al., 2023). Given the critical challenges of managing wounds in trauma patients, fibronectin may significantly help wound healing. During the healing process, fibronectin facilitates the spread of platelets at the injury site and supports the movement and attachment of cells into the wound area (Grinnell, 1984). Fibronectin enhances phagocytosis, the process of ingesting foreign pathogens, which plays a key role in sterilizing wound beds throughout the healing process (Grinnell, 1984). Fibronectins have specific regions that help bond to collagen and cells, like fibroblasts and macrophages, fibrin, and certain sugars, aiding in the adhesion of collagen and cells (Figure 4; Landén et al., 2016; Clark et al., 2018).
| Figure 4. Fibronectin enhances the integrity of the extracellular matrix. |
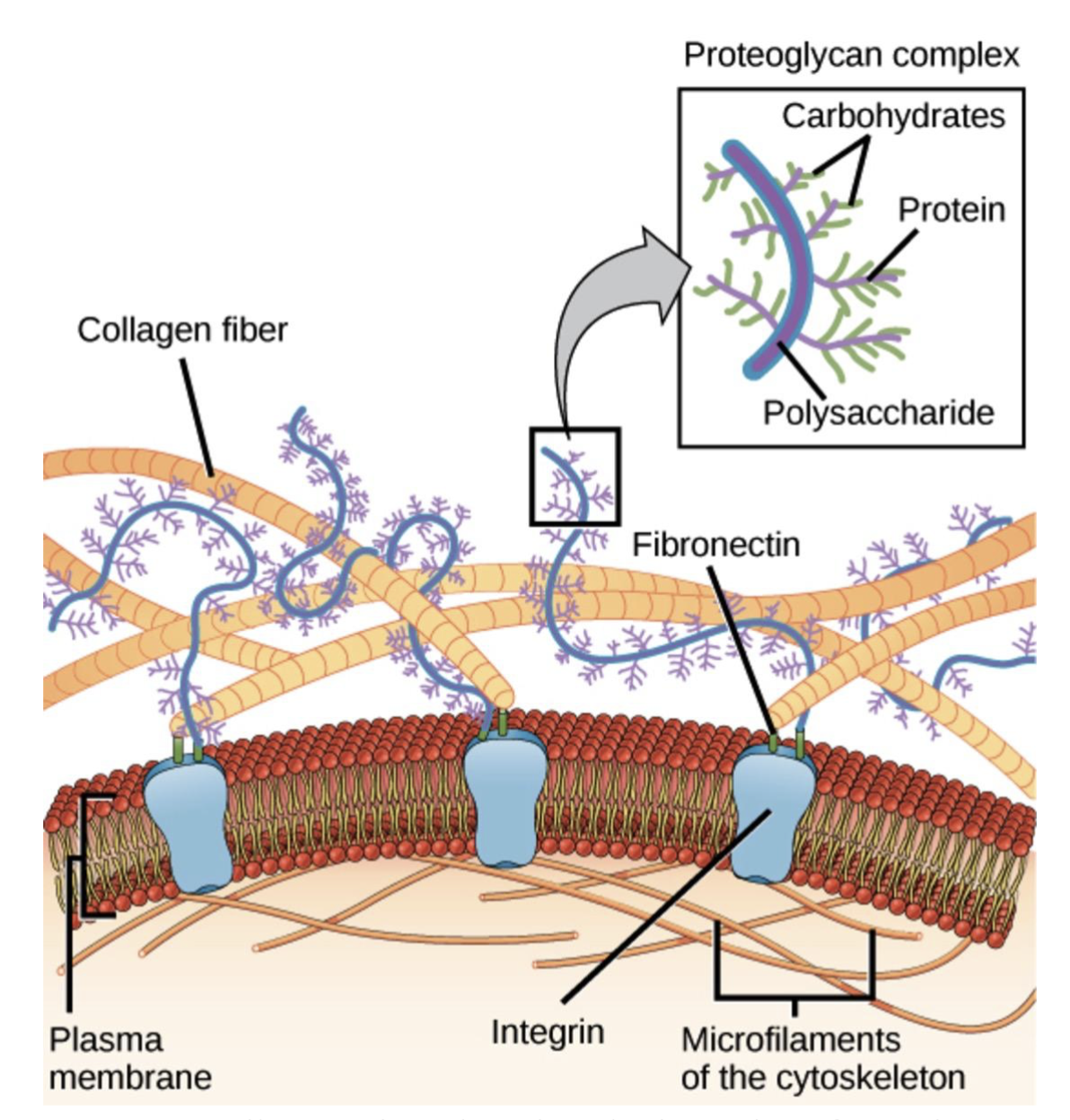
|
This diagram shows how integrin transmembrane receptors connect the ECM to intracellular cytoskeletal microfilaments. Fibronectin links collagen fibers and proteoglycans and facilitates cell-ECM adhesion. These interactions are crucial for the wound-healing process (Clark, Douglas, & Choi, 2018).
Fibrin plays a crucial role in maintaining hemostasis through the formation of a clot. It catalyzes the coagulation process and increases the strength of platelet clots (Figure 5; Gorane, 2023). Fibrin is formed from the long fibrous chain fibrinogen, a protein made by the liver and found in blood plasma.
Both fibronectin and fibrin contribute to all four phases of wound healing: hemostasis, inflammation, proliferation, and remodeling, particularly in the proliferation phase (Trinh et al., 2022). When an acute injury occurs, prothrombinase first activates prothrombin to thrombin. Then, thrombin cuts fibrinogen into fibrin monomers (Goh et al., 2016; Krishnaswamy, 2013). Then, fibrin monomers form fibrin strands through a process of polymerization. Long fibrin strands entangle the platelet to create a spongy mass that gradually hardens and contracts to form a blood clot (Britannica, Fibrin). Additionally, fibrinogen helps modulate inflammatory cellular reactions and increases plasma concentration after injury, facilitating inflammation and cell proliferation (Levy et al., 2012).
| Figure 5. Thrombin cleaves fibrinogen into fibrin during the wound healing process. The process begins with the release of clotting factors, leading to the conversion of prothrombin to thrombin. Thrombin then catalyzes the transformation of soluble fibrinogen into insoluble fibrin strands. These strands interact with platelets at the injury sites to form a blood clot (Gorane, 2023; Cleveland Clinic, 2024). |
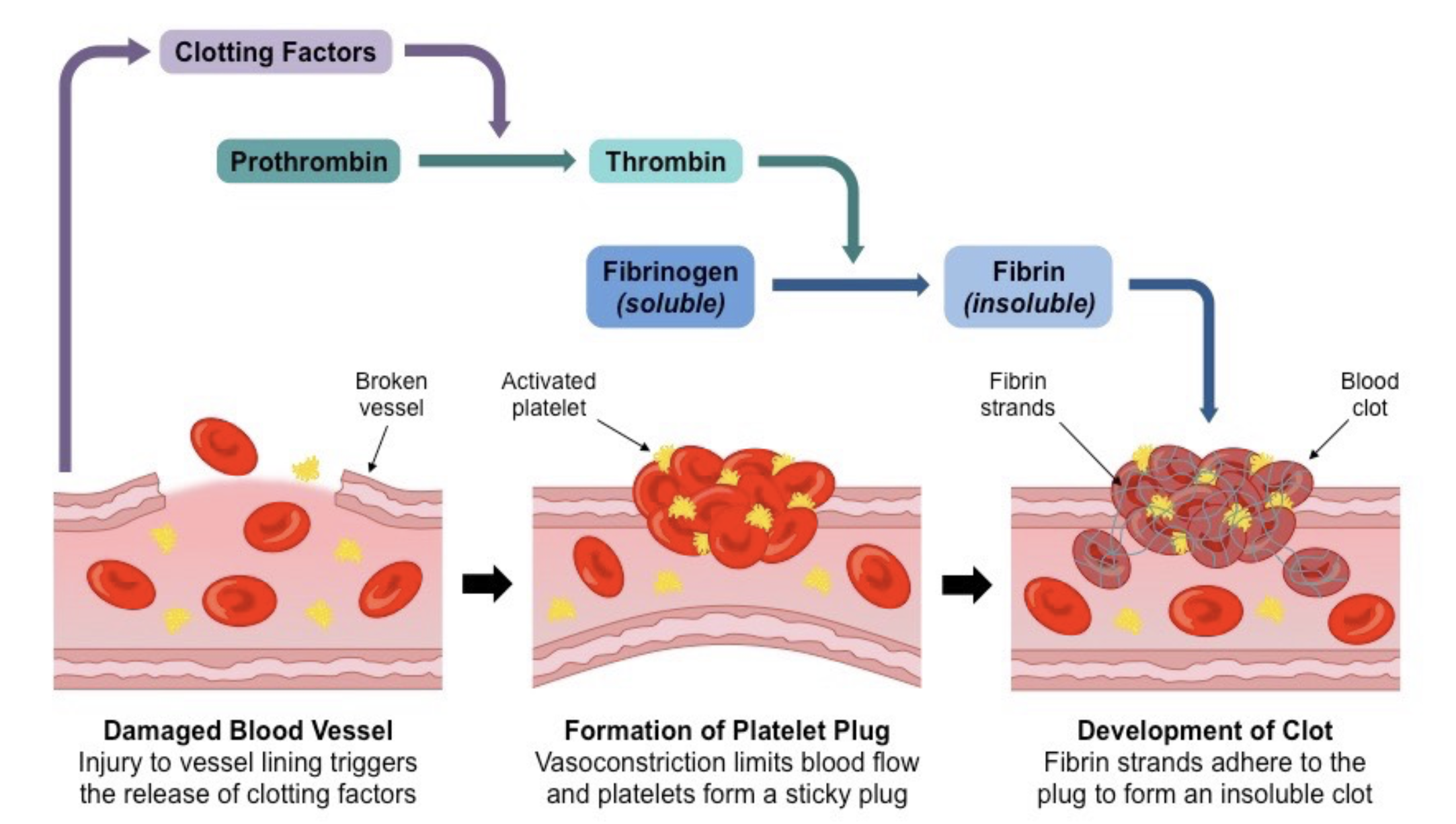
|
In the proliferation phase, the initial wound covering is replaced by granulation tissue comprising fibroblasts, immune cells, blood vessels, and collagen. This tissue helps restore some structure and function to the damaged skin (Landén et al., 2016).
Beyond that, recent studies have highlighted the critical role of fibronectin beneath the migrating outer skin tissues during re-epithelialization, the process of covering a wound with new skin cells during the proliferation phase (Lenselink, 2013; Rousselle et al., 2019; Tan & Dosan, 2019). This process is supported by clot formation and the migration of new skin cells into the wound (Larjava et al., 2000-2013). When endothelial cells and fibroblasts enter the wound bed, they deposit fibronectin, essential for granulation tissue formation (Lenselink, 2013).
In complex procedures like skin grafts and transplants, fibronectin boosts wound healing, enhances immune response by promoting phagocytosis, and maintains a sterile wound bed. Fibronectin also stimulates angiogenesis, improving nutrient and growth factor delivery. This innovative treatment ultimately provides safer and more effective wound care for patients.
Additionally, fibronectin production helps guide the epithelial cells toward the injury site (Larjava et al., 2000-2013; Rousselle et al., 2019). Capillary sprouts involved in angiogenesis (the process of forming new blood vessels) also specifically invade the fibrin/fibronectin-rich clot and organize it into a microvascular network within the granulation tissue (Tonnesen et al., 2000).
Emerging fibronectin-based therapies enhance wound healing, particularly in challenging cases such as chronic wounds and diabetic ulcers. Studies suggest that fibronectin and fibrin, combined with growth factors, may accelerate the healing of complex wounds (Patten et al., 2020). Especially, the topical delivery of fibronectin significantly improves the wound healing of the radiation-damaged skin, in which fibronectin expression is significantly and consistently downregulated (Johnson et al., 2017).
The sustainable bacterial fibronectin and fibrin provide a reliable, high-yield source for surgical procedures involving exposed wounds. Usually, fibrin is the first coagulation factor that decreases to a critically low level (Levy et al., 2012). Processed fibrin strands accelerate wound healing and clot formation without requiring additional thrombin cleavage from the body.
Systems Level
The aim of this project is to engineer E. coli for the production of recombinant fibronectin and fibrin strands, which could subsequently be applied to enhance existing wound healing technologies and ultimately improve the treatment of open wounds. We will use the plasmid pET28-H12TEV and transform it into the BL21 strain of E. coli (Figure 6; Ma, n.d.; Addgene Plasmid #208635). In order to achieve high yield of the three proteins, the expression vector comprises of the deop1p2 promoter, SOD (human superoxide dismutase) gene connected to the FN1 (fibronectin) gene, F2 (prothrombin) gene, F5 (coagulation factor V) gene, F10 (coagulation factor X) gene, FGA, FGB, FGG (fibrinogen) gene set, GFP (green fluorescent protein) gene, and the trpA terminator will be added in the plasmid (Figure 7). GFP facilitates the easy monitoring of gene expression levels, enabling further investigation and optimization. Additionally, GST (Glutathione-S-transferase enzyme) tags will be added to genes encoding fibronectin and fibrinogen production. After plasmid introduction via the calcium chloride transformation protocol, the bacteria will be incubated at 37°C. During cultivation, the activated thrombin cleaves the produced fibrinogen into fibrin monomers, which subsequently will polymerize into fibrin. Affinity chromatography, a protein filtration system, will then be used to remove thrombin, coagulation factors, and cellular debris, leaving behind fibronectin and polymerized fibrin as the final product. Our choices for constructing the ECM protein-yielding system are supported by previous research, which demonstrates compatibility and positive results, thereby supporting the system’s feasibility (Gogoi et al., 2006).
| Figure 6. Backbone pET28-H12TEV Plasmid with Original Components. This pET28-H12TEV plasmid map illustrates the origin of replication, the Kanamycin resistance sequence, the Lactose Operon, the repressor of primers (ROP), and the T7 terminator, which is compatible with E. coli for protein production. |
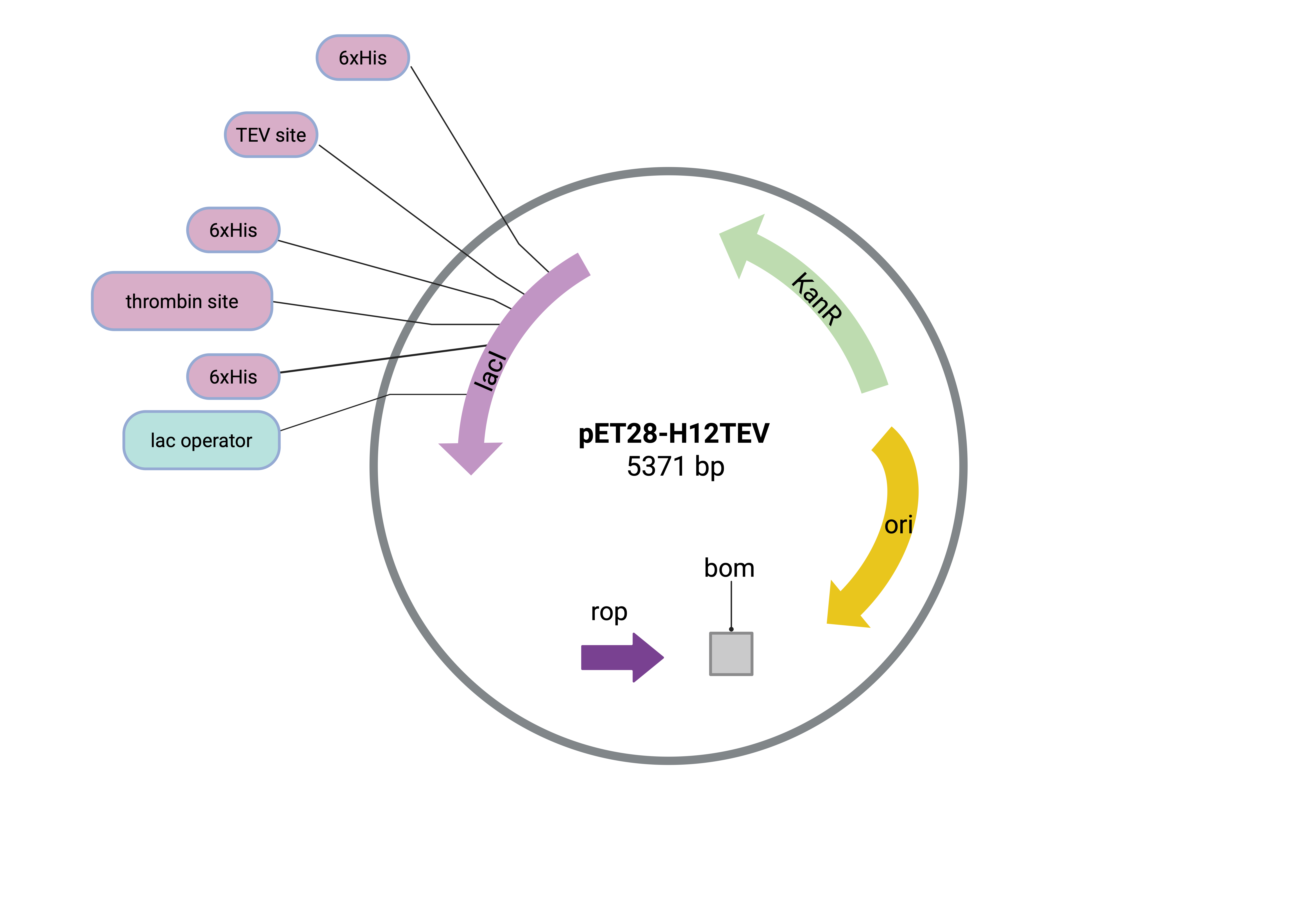
|
| Figure 7. Plasmid map of pETFN-OI666, engineered from pET28-H12TEV. |
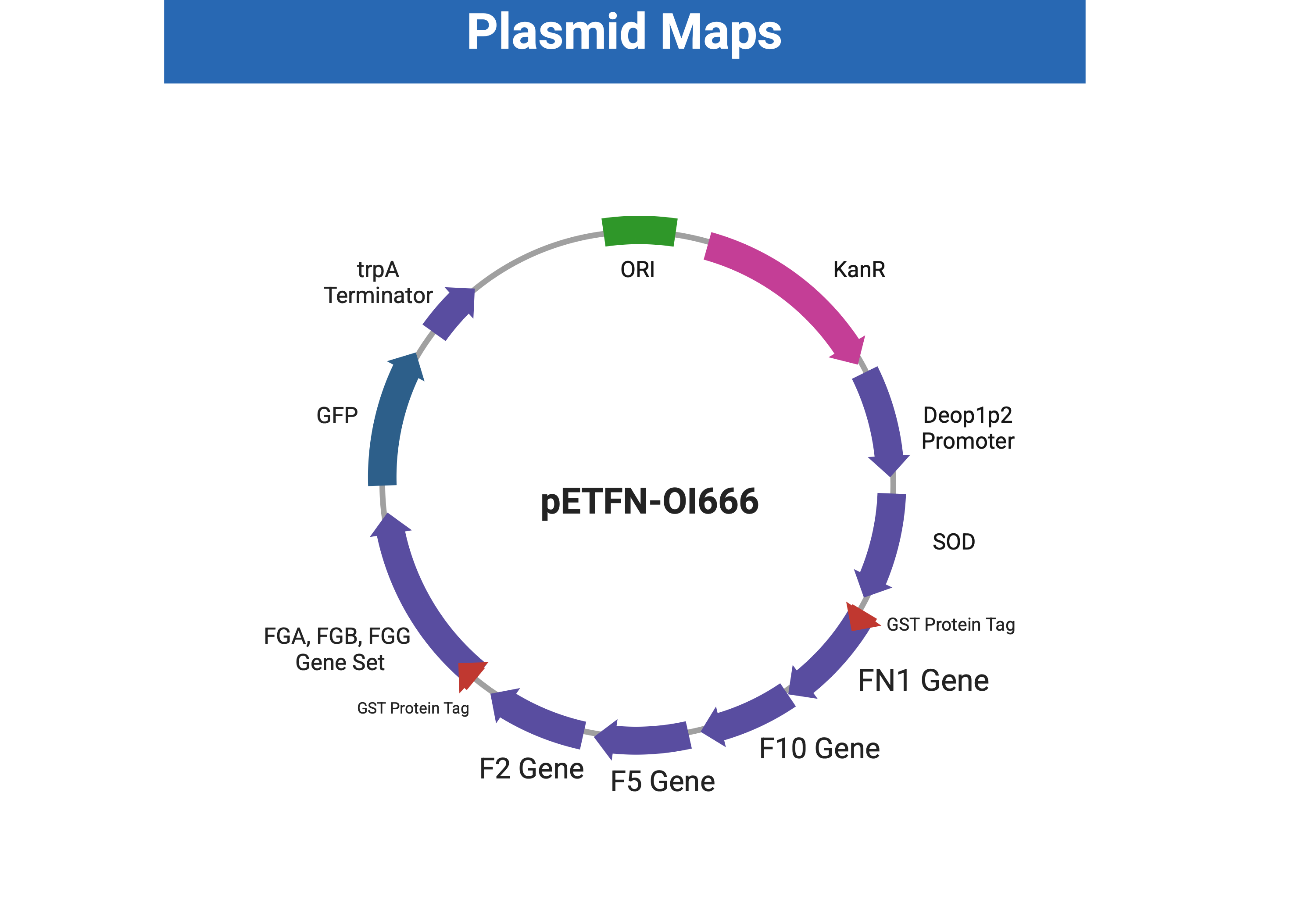
|
This plasmid is a prototype for fibronectin expression, containing tagged FN1, F2, F5, F10, and tagged FGA, FGB, FGG expression for easier purification. KanR (Kanamycin resistance) is used as a selectable marker, and GFP serves as a signal for the success of the production process. The expression is initiated by the deop1p2 promoter and stopped by the trpA terminator.
Device Level
The construction of the plasmid pETFN-OI666 involves the manipulation of the backbone plasmid pET28-H12TEV, codon optimization of the targeted DNA, and insertion of the entire expression vector.
We plan to remove the ROP section and the thrombin cleavage site from plasmid pET28-H12TEV by PCR amplification with opposite-direction primers that leave out the unwanted region and re-ligate the plasmid to circularize it. The ROP protein keeps a low copy number of plasmid in E. coli.
Since FN1, F2, F5, F10, and FGA, FGB, and FGG gene sets are stranded directly from Homo Sapiens, eukaryotic species, these sequences may contain a large number of “rare codons” that are infrequently used in E. coli, including arginine (AGA, AGG), isoleucine (AUA), leucine (CUA), and proline (CCC). The incompatibility may result in a decrease or elimination of the expression of the targeted protein. By undergoing codon optimization, the rare codons in the FN1, F2, F5, F10, and FGA, FGB, and FGG gene sets will be replaced with codons more common in E. coli.
FN1 and FGA, FGB, and FGG gene sets are tagged with Glutathione-S-Transferase (GST) through PCR with primers containing the GST sequence for the later purification process. The whole gene sequence comprises the deop1p2 promoter, the SOD gene connected with the tagged FN1 gene, the F2 gene, the F5 gene, the F10 gene, the tagged FGA, FGB, and FGG gene set, the GFP gene, and the trpA terminator. These genes are inserted into the altered pET28-H12TEV backbone plasmid to form the end product pETFN-OI666 through ligation.
Parts Level
E. coli has long been used for recombinant protein production due to its fast growth, easy manipulation, and cost‐effectiveness. The BL21(DE3) strain of E. coli stands out among other cell lines as a host for recombinant proteins. Its doubling time is around 20 minutes in rich medium. It grows faster than lines such as K-12 in a minimal medium, partly attributed to its absence of flagella and flagella biosynthesis (an energy-intensive protein synthesis process). E. coli B’s deficiency in protease (an enzyme that breaks down protein) ensures a longer lifetime for recombinant protein and more protein yield during purification, since cell disruption causes a massive release of its protease. Furthermore, E. coli B secretes more protein than other laboratory strains through its second type II secretion (T2S) system, thereby increasing protein production (Rosano et al., 2019).
Plasmid pET28-H12TEV has proven effective for large-sized protein expression and is highly compatible with the E. coli strain BL21. It contains a large variety of restriction sites that ease the manipulation and extension. The lactose operon enables E. coli to metabolize lactose when glucose, the preferred fuel, is absent (Tankeshwar, 2019). Also, the reaction between beta-galactosidase in the lactose operon and X-gal in the growth medium provides an alternative selection method. The beta-galactosidase enzyme hydrolyzes X-gal producing a blue product that causes the E. coli colonies to turn blue (Juers et al., 2012). The lower price of lactose lowers the overall cost of this production system, allowing the protein to be produced at a lower cost and making the product more accessible to underserved areas. The expression vector is composed of the deop1p2 promoter, the SOD gene connected with the targeted protein gene, and the trpA terminator. Implementing this expression vector system for human insulin production has proven successful (Mikiewicz et al., 2017). The deop1p2 promoter is a constitutive promoter that drives continuous gene expression regardless of external or internal cellular changes (Gene Regulation and Constitutive Promoters in Biotechnology (GRCPB), 2025). SOD acts as a signal peptide to enhance protein expression, overcoming the relatively low plasmid copy number due to the high metabolic burden associated with large protein expression.(Mikiewicz et al., 2017). Our targeted proteins include the GST-tagged FN1 gene, which encodes fibronectin, and the GST-tagged FGA, FGB, and FGG genes, which encode fibrinogen. GST-tag enhances soluble expression and facilitates protein purification during later affinity chromatography. We also insert the F2 gene, which encodes prothrombin, and the F5 and F10 genes, which encode coagulation factor V and X, respectively. These genes combine to form prothrombinase. Through the activation by prothrombinase, prothrombin is converted to thrombin, which subsequently cleaves fibrinogen and initiates fibrin polymerization (Krishnaswamy, 2013). GFP allows easy monitoring of gene expression levels for further investigation and adjustment. The GFP fluorescence in the plasmid reflects the successful production of protein.
Safety
Our project adheres to standard biosafety practices and aims to minimize risks to humans or the environment. We will work exclusively with a non-pathogenic E. coli strain BL21(DE3), a Biosafety Level 1 (BSL-1) organism recognized as safe for laboratory use (Thermo Fisher Scientific (TFS), 2016). All experiments will be conducted in a BSL-1 certified laboratory to prevent environmental release and minimize biosafety risks. Throughout the experiment, all steps must follow the protocols associated with the kit, and personal protective equipment should be worn at all times.
Prior to harvesting the ECM proteins, E. coli will be neutralized by a freeze-thaw cycle, which can lead to frostbite if the standard safety precautions are not followed. Additionally, fibrinogen and fibronectin will be filtered with our filtration system before being added to open wounds, leaving only ECM proteins, which are safe for open wound treatments.
In affinity chromatography, standard wash buffer components include the use of Phosphate-buffered saline (PBS), and the addition of 0.5 mL of Tagmentase Dilution Buffer (TNGT) elution buffer. These buffers will elute the GST-tagged protein (Figure 8; Smith & Johnson, 2014). To prevent endotoxin contamination, we will use Pierce™ Rapid Gel Clot Endotoxin Assay Kits for endotoxin detection (TFS, n.d.). If endotoxin is detected, we will use Pierce™ High-Capacity Endotoxin Removal Resin (Thermo Fisher Scientific [TFS], n.d.). TNGT elution buffer consists of chemicals with low toxicity under laboratory conditions; however, proper safety precautions must be strictly followed, and safety equipment must be worn (TFS, 2021; TFS, 2023).
| Figure 8. Affinity Chromatography Filtration System for GST-tagged Protein. Affinity chromatography filters unwanted proteins and cell debris through a filtration column. Fibrin and Fibronectin are bound to the MagneGST GSH-particle by their GST tag, which is later eluted by TNGT elution buffer (Thermo Fisher Scientific, n.d.). |
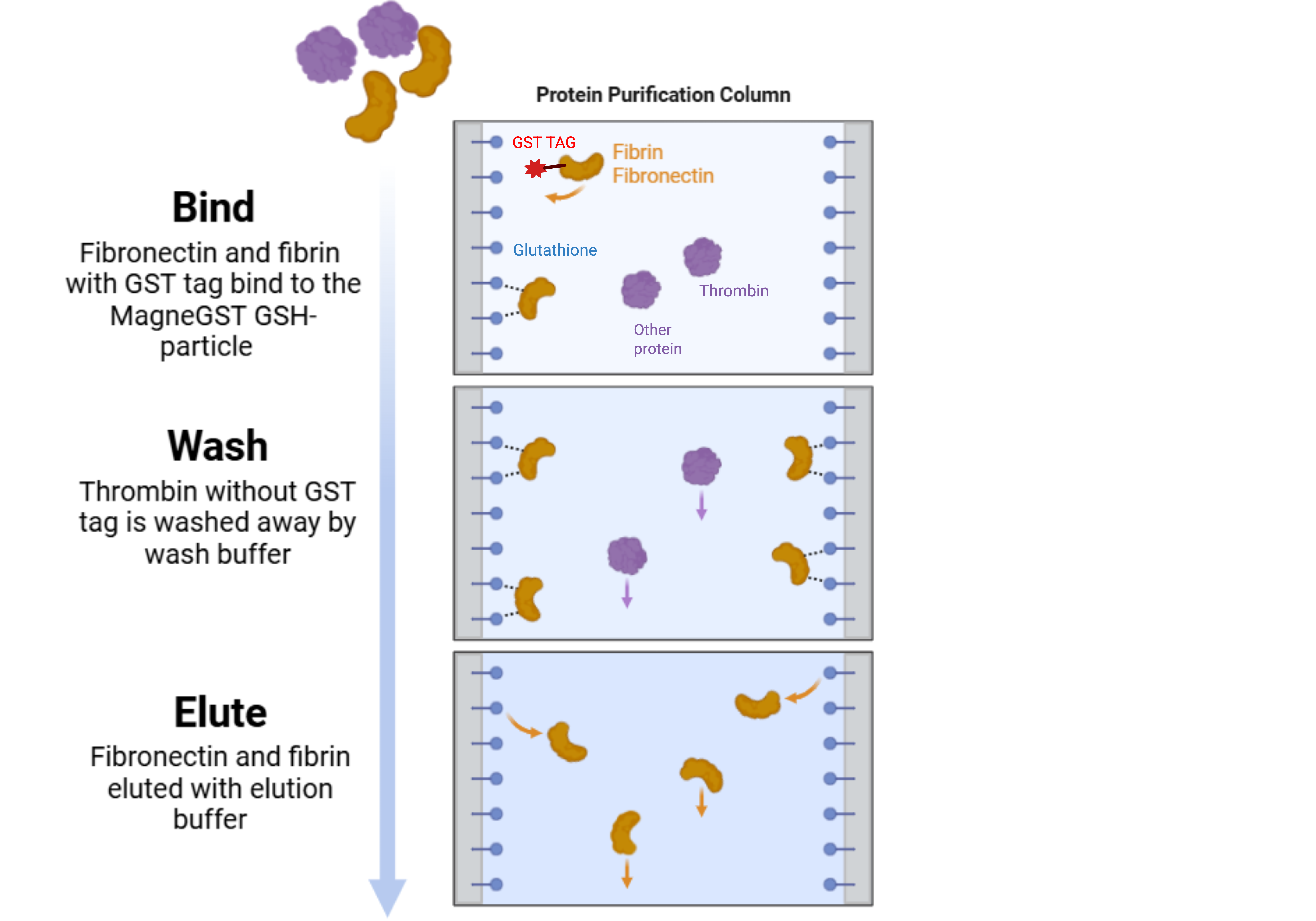
|
Discussions
This project aims to develop a sustainable and efficient system for producing fibronectin and fibrin, thereby enhancing wound healing. Using E. coli BL21 as the chassis, we introduce the engineered pETFN-OI666 plasmid, which encodes fibronectin, prothrombin, and fibrinogen. This design mimics the human pathway of fibronectin production, involving thrombin activation, cleavage of fibrinogen, and fibrin polymerization to ensure the effective production of an end product.
Fibronectin and fibrin are essential for wound healing, but are quickly lost through bleeding. Their slow natural replenishment increases the risk of infection and blood loss. To address this, we propose harnessing fast-growing microorganisms to produce these proteins at high yields in a sustainable manner. The recombinant proteins supplement the body’s healing response, accelerating recovery and reducing associated risks.
After evaluating various chassis for protein production, we identified E. coli as the optimal choice due to its ease of manipulation, rapid growth, and cost-effectiveness. Specifically, the BL21 strain was selected for its short doubling time, low metabolic burden, and high protein yield (Rosano et al., 2019). Protein expression in E. coli is a well-established technology, and its proven ability to produce human proteins, such as insulin, demonstrates its suitability for our project despite concerns about cross-species gene compatibility.
The engineered pETFN-OI666 comprises several elements that maintain the sustainability of the protein expression, augment the productivity, and ease the selection and monitoring process. It encodes all three proteins together, ensuring a higher efficiency. The GST-tag enhances the yield of soluble proteins, thereby improving the quality of the final product after protein purification. The kanamycin selection mark helps to select the wanted bacteria with the transformed plasmid, and the GFP signals the gene transcription and translation process.
However, several limitations and uncertainties may impact production efficiency and final product quality. These include codon optimization levels, the absence of the ROP protein, unknown metabolic burden on E. coli, and concerns around protein solubility and application safety.
Studies have shown the unanticipated effects of codon optimization. Codon changes may affect protein conformation and stability. Mutations and site changes in post-translational modification may alter the protein’s function (Mauro & Chappell, 2014). Determining the optimal level of codon optimization for maximal expression and solubility requires further validation. We plan to assess gene expression using electrophoresis and quantitative PCR (qPCR) with custom primers.
The ROP gene regulates the copy number of the backbone plasmid (pET28-H12TEV), which helps maintain a constant copy number (Castagnoli et al., 1989). In our design, ROP is removed to increase plasmid copy number; however, its absence may affect plasmid stability, E. coli growth, and protein yield. Comparing ROP-containing and ROP-deficient plasmids throughout the production process is crucial for evaluating sustainability and optimizing performance.
Human proteins are significantly larger and more complex than proteins in E. coli (BioNumbers, n.d.). Studies have shown that the probability of soluble protein expression decreases with increasing molecular weight, especially for proteins exceeding 60 kDa (Francis & Page, 2010). Expressing 3 human proteins within one expression vector may pose a high metabolic burden and decrease soluble expression, as the total molecular weight of targeted proteins exceeds 60 kDa (230- 270 kDa for fibronectin, 72kDa for prothrombin, 340 kDa for fibrinogen) (Dalton & Lemmon, 2021; Lakna, 2023; Tiscia & Margaglione, 2018). Close monitoring for E. coli growth and protein function testing is crucial to examine the feasibility of our design.
Before applying the recombinant proteins in human wound healing, their properties must be thoroughly evaluated, including potential mutations, functionality, toxicity, and immune responses. Although a previous study demonstrates the feasibility of topical fibronectin use, the viability of topical delivery for both fibronectin and fibrin to wound sites requires further evaluation (Johnson et al., 2017). Under surgical conditions, the blood clotting ability of the end product may become a factor in thrombus formation. In vitro analyses and in vivo testing using lab mice and organoids will help simulate real-world performance to verify the topical delivery method for open wound sites, as well as explore a safer in vivo delivery method for surgical conditions. Comparative studies of proteins produced by E. coli with different plasmid designs will identify the optimal construct and guide future improvements.
Next Steps
Since our project introduces a novel approach to medicated bandages, we need to test and quantify the fibrinogen and fibronectin produced. First, we will use a calcium chloride transformation protocol to insert the pETFN-OI666 plasmid, containing FN1, F2, FGA, FGB, and FGG genes, into E. coli strain BL21.
Next, we will have to identify the optimal culture time for the bacteria. The transformed bacteria will be incubated at 37°C for 24 hours for the appearance of colonies on agar plates, then verified for successful transformation using fluorescence microscopy to detect GFP (“Incubation Time”, n.d.; Wei et al., 2014). Afterwards, successfully transformed bacteria will be isolated through kanamycin selection. In order to obtain quantitative data, Western Blotting will be used to quantify the presence of the ECM proteins produced by the bacterial culture (Ha et al., 2023).
Once a successful transformation is verified via fluorescence microscopy, and protein production is verified via Western Blotting, we will proceed with optimization and protein filtration. This phase includes optimizing incubation time by culturing bacteria for different durations and measuring protein yield, lysing the bacteria using freeze-thaw cycles followed by centrifugation to separate proteins from cell debris, and purifying the proteins through affinity chromatography. The filtration column will be washed with PBS and eluted with TNGT buffer to isolate ECM proteins (TFS, n.d.). To measure protein concentration, we will use a UV-Vis spectrophotometer to measure the absorbance at 280 nm (Peak Proteins, n.d.). In the future, in vitro studies using the C57BL/6 mouse model, commonly employed for wound healing testing y, will be conducted to further evaluate the effectiveness of our bandage treatment (van den Borne et al., 2009).
Author Contributions
X.Y conceived the project idea. Z.C., C.C., R.W., and X.Y. contributed to the research and manuscript preparation. C.C. and R.W. created the images and video. Z.C., C.C., R.W., and X.Y. completed the citations and worked on proofreading.
Acknowledgements
We are very grateful to Western Reserve Academy for providing this opportunity for us to proceed with our design. We also extend our appreciation to Dr. Pethel and Mr. Switzer for guiding us through the entire construction and paper-writing process. Their expertise in synthetic biology contributes to the accuracy and viability of our design. Their endeavor and mentorship are crucial for deepening our comprehension of synthetic biology through this design process.
References
BioNumbers. (n.d.). Average protein size (BNID 113349). Retrieved May 6, 2025, from https://bionumbers.hms.harvard.edu/bionumber.aspx?id=113349
Castagnoli L, Scarpa M, Kokkinidis M, Banner DW, Tsernoglou D, Cesareni G. Genetic and structural analysis of the ColE1 Rop (Rom) protein. EMBO J. 1989 Feb;8(2):621-629. doi: 10.1002/j.1460-2075.1989.tb03417.x. PMID: 2721494; PMCID: PMC400849.
Centers for Disease Control and Prevention. (2021, December 2). Economic cost of injury – United States, 2019. Centers for Disease Control and Prevention. https://www.cdc.gov/mmwr/volumes/70/wr/mm7048a1.htm
Centers for Disease Control and Prevention. (2024, July 26). FastStats – accidents or unintentional injuries. Centers for Disease Control and Prevention. https://www.cdc.gov/nchs/fastats/accidental-injury.htm
Clark, M. A., Douglas, M., & Choi, J. (2018, March 28). 4.6 Connections between Cells and Cellular Activities – Biology 2e | OpenStax. https://openstax.org/books/biology-2e/pages/4-6-connections-between-cells-and-cellular-activities
Cleveland Clinic. (2024, April 24). Hemorrhage. Cleveland Clinic. https://my.clevelandclinic.org/health/diseases/hemorrhage
Cleveland Clinic. (2024, May 1). Contact dermatitis: Symptoms, causes, types & treatments. Cleveland Clinic. Retrieved November 18, 2024, from https://my.clevelandclinic.org/health/diseases/6173-contact-dermatitis
Dalton, C. J., & Lemmon, C. A. (2021). Fibronectin: Molecular Structure, Fibrillar Structure and Mechanochemical Signaling. Cells, 10(9), 2443. https://doi.org/10.3390/cells10092443
Encyclopædia Britannica, Inc. (n.d.). Fibrin. Encyclopædia Britannica. https://www.britannica.com/science/fibrin
Epidermis (outer layer of skin): Layers, function, structure. (2024, May 1). Cleveland
Clinic. Retrieved November 18, 2024, from https://my.clevelandclinic.org/health/body/21901-epidermis
Fallah, N., Rasouli, M. & Amini, M.R. (2021). The current and advanced therapeutic modalities for wound healing management. Journal of Diabetes Metabolic Disorders 20, 1883–1899 (2021). https://doi.org/10.1007/s40200-021-00868-2
Francis, D. M., & Page, R. (2010). Strategies to optimize protein expression in E. coli. Current protocols in protein science, Chapter 5(1), 5.24.1–5.24.29. https://doi.org/10.1002/0471140864.ps0524s61
Gene regulation and constitutive promoters in biotechnology. (2025, April 30). BiologyInsights.https://biologyinsights.com/gene-regulation-and-constitutive-promoters-in-biotechnology/
Gentamicin side effects: Common, severe, long-term. (2024, July 21). Drugs.com. Retrieved November 18, 2024, from https://www.drugs.com/sfx/gentamicin-side-effects.html
Goh, E., Kirby, G., Jayakumar, R., Liang, X.-J., & Tan, A. (2016). Accelerated wound healing using nanoparticles. Nanoscience in Dermatology (pp. 287-306).
Gogoi SK, Gopinath P, Paul A, Ramesh A, Ghosh SS, Chattopadhyay A. (2006) Green fluorescent protein-expressing Escherichia coli as a model system for investigating the antimicrobial activities of silver nanoparticles. Langmuir,22(22):9322-9328. doi: 10.1021/la060661v.
Gorane, J. (2023, November 9). Blood coagulation (Blood clotting). CBSE Class Notes Online Classnotes123. https://classnotes123.com/blood-coagulation-blood-clotting/Hemorrhage. Cleveland Clinic. (2024, November 8). https://my.clevelandclinic.org/health/diseases/hemorrhage
Grinnell, F. (1984). Fibronectin and wound healing. Journal of Cellular Biochemistry, 26(2), 107-116. https://doi.org/10.1002/jcb.240260206
Ha, B. G., Jang, Y.-J., Lee, E., Kim, B.-G., Myung, K., Sun, W., & Jeong, S.-J. (2023). Isolation and identification of extracellular matrix proteins from oil-based casperized mouse brains for matrisomal analysis. Heliyon, 9(4), e14777. https://doi.org/10.1016/j.heliyon.2023.e14777
Hemostasis. (n.d.). Cleveland Clinic. Retrieved November 17, 2024, from https://my.clevelandclinic.org/health/symptoms/21999-hemostasis
Incubation time. (n.d.). ScienceDirect Topics. Retrieved May 6, 2025, from https://www.sciencedirect.com/topics/biochemistry-genetics-and-molecular-biology/incubation-time
Johnson, M. B., Pang, B., Gardner, D. J., Niknam-Benia, S., Soundarajan, V., Bramos, A., Perrault, D. P., Banks, K., Lee, G. K., Baker, R. Y., Kim, G. H., Lee, S., Chai, Y., Chen, M., Li, W., Kwong, L., Hong, Y. K., & Wong, A. K. (2017). Topical fibronectin improves wound healing of irradiated Skin. Scientific reports, 7(1), 3876. https://doi.org/10.1038/s41598-017-03614-y
Juers, D. H., Matthews, B. W., & Huber, R. E. (2012). LacZ β-galactosidase: structure and function of an enzyme of historical and molecular biological importance. Protein science: a publication of the Protein Society, 21(12), 1792–1807. https://doi.org/10.1002/pro.2165
Kolimi, P., Narala, S., Nyavanandi, D., Youssef, A. A. A., & Dudhipala, N. (2022). Innovative Treatment Strategies to Accelerate Wound Healing: Trajectory and Recent Advancements. Cells, 11(15), 2439. https://doi.org/10.3390/cells11152439
Krishnaswamy S. (2013). The transition of prothrombin to thrombin. Journal of thrombosis and haemostasis11 (Suppl 1), 265–276. https://doi.org/10.1111/jth.12217
Lakna. (2023, September 21). What is the difference between thrombin and prothrombin Pediaa.com. https://pediaa.com/what-is-the-difference-between-thrombin-and-prothrombin/
Landén, N. X., Li, D., & Ståhle, M. (2016). Transition from inflammation to proliferation: A critical step during wound healing. Cellular and Molecular Life Sciences, 73(20), 3861-3885. https://doi.org/10.1007/s00018-016-2268-0
Larjava, H., Koivisto, L., & Häkkinen, L. (n.d.). Keratinocyte Interactions with Fibronectin during Wound Healing. In Madame Curie Bioscience Database [Internet]. https://www.ncbi.nlm.nih.gov/books/NBK6391/
Lenselink, E. A. (2013). Role of fibronectin in normal wound healing. International Wound Journal, 12(3), 313-316. https://doi.org/10.1111/iwj.12109
Levy, J. H., Szlam, F., Tanaka, K. A., & Sniecienski, R. M. (2012). ibrinogen and hemostasis: A primary hemostatic target for the management of acquired bleeding. Anesthesia & Analgesia, 114(2), 261–274. https://doi.org/10.1213/ANE.0b013e31822e1853
Ma, Q. (n.d.). PET28-H12TEV. Addgene. Retrieved December 6, 2024, from https://www.addgene.org/208635/
Markiewicz-Gospodarek, A., Kozioł, M., Tobiasz, M., Baj, J., Radzikowska-Büchner, E., & Przekora, A. (2022). Burn wound healing: Clinical complications, medical care, treatment, and dressing types: The current state of knowledge for clinical practice. International Journal of Environmental Research and Public Health, 19(3), 1338. https://doi.org/10.3390/ijerph19031338
Mauro, V. P., & Chappell, S. A. (2014). A critical analysis of codon optimization in human therapeutics. Trends in molecular medicine, 20(11), 604–613. https://doi.org/10.1016/j.molmed.2014.09.003
Mikiewicz D, Bierczyńska-Krzysik A, Sobolewska A, Stadnik D, Bogiel M, Pawłowska M, Wójtowicz-Krawiec A, Baran PA, Łukasiewicz N, Romanik-Chruścielewska A, Sokołowska I, Stadnik J, Borowicz P, Płucienniczak G, Płucienniczak A. (2017) Soluble insulin analogs combining rapid- and long-acting hypoglycemic properties – From an efficient E. coli expression system to a pharmaceutical formulation. PLoS ONE, ,e0172600. https://doi.org/10.1371/journal.pone.0172600
Nunez, K. (2019, February 13). Debridement: Types, Recovery, Complications & More. Healthline. https://www.healthline.com/health/debridement#when-its-done
Patten, J., & Wang, K. (2020, September 19). Fibronectin in development and wound healing. Advanced Drug Delivery Reviews. https://www.sciencedirect.com/science/article/pii/S0169409X20301290?via%3Dihub
Peak Proteins. (n.d.). Protein concentration by UV absorbance at 280 nm280nm. Peak Proteins. https://peakproteins.com/wp-content/uploads/2022/06/PPP_Protein-Conc-by-UV.pdf
Potts, Carla. (2016) Statement of non-pathogenicity. Thermo Fisher Scientific.https://assets.thermofisher.com/TFS-Assets/LSG/manuals/PG1572-PJT1331-COL12226-Statement-NonPathogenicity-FHR.pdf
Rosano, G. L., Morales, E. S., & Ceccarelli, E. A. (2019). New tools for recombinant protein production in Escherichia coli: A 5-year update. Protein science, 28(8), 1412–1422. https://doi.org/10.1002/pro.3668
Rousselle, P., Braye, F., & Dayan, G. (2019). Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Advanced Drug Delivery Reviews, 146, 344-365. https://doi.org/10.1016/j.addr.2018.06.019
Sicari, B. M., Londono, R., Dziki, J. L., & Badylak, S. F. (2023). Extracellular matrix as a bioscaffold for tissue engineering. Tissue Engineering, 137-172. https://doi.org/10.1016/b978-0-12-824459-3.00005-6
Smith, D. B., & Johnson, K. S. (2014). Purification of GST-tagged proteins. Methods in Enzymology, 529, 257–280. https://doi.org/10.1016/B978-0-12-397863-9.00016-2
Tan, S., & Dosan, R. (2019). Lessons From Epithelialization: The Reason Behind Moist Wound Environment. The Open Dermatology Journal, 13, 34-40. https://doi.org/10.2174/1874372201913010034.
Tankeshwar, A. (2019, November 27). Lac Operon: Mechanism and Regulation. Learn Microbiology Online. https://microbeonline.com/lac-operon-mechanism/
Thermo Fisher Scientific. (2021, December 24). Safety data sheet: Tris(hydroxymethyl)aminomethane hydrochloride (Revision No. 7). https://www.fishersci.com
Thermo Fisher Scientific. (2021, December 24). Safety data sheet: Tween® 20 (Revision No.7). https://www.fishersci.com
Thermo Fisher Scientific. (2023, October 13). Safety data sheet: Sodium chloride (Revision No. 8). https://www.fishersci.com
Thermo Fisher Scientific. (2021, December 24). Safety data sheet: Glutathione, reducing (Revision No. 5). https://www.fishersci.com
Thermo Fisher Scientific. (n.d.). Overview of affinity purification. Thermo Fisher Scientific. https://www.thermofisher.com/us/en/home/life-science/protein-biology/protein-biology-learning-center/protein-biology-resource-library/pierce-protein-methods/overview-affinity-purification.html
Thermo Fisher Scientific. (n.d.). PureLink™ RNA Mini Kit. Thermo Fisher Scientific. https://www.thermofisher.com/order/catalog/product/A43879
Thermo Fisher Scientific. (n.d.). Pierce™ BCA Protein Assay Kit. Thermo Fisher Scientific. https://www.thermofisher.com/order/catalog/product/88270
Tiscia, G. L., & Margaglione, M. (2018). Human Fibrinogen: Molecular and genetic aspects of congenital disorders. International journal of molecular sciences, 19(6), 1597. https://doi.org/10.3390/ijms19061597
Tonnesen, M. G., Feng, X., & Clark, R. A.F. (2000). Angiogenesis in wound healing. Journal of Investigative Dermatology Symposium Proceedings, 5(1), 40-46. https://doi.org/10.1046/j.1087-0024.2000.00014.x
Tribble, D. R., Krauss, M. R., Murray, C. K., Warkentien, T. E., Lloyd, B. A., Ganesan, A., Greenberg, L., Xu, J., Li, P., Carson, M. L., Bradley, W., & Weintrob, A. C. (2018). Epidemiology of trauma-related infections among a combat casualty cohort after initial hospitalization: The trauma infectious disease outcomes study. Surgical Infections, 19(5), 494–503. https://doi.org/10.1089/sur.2017.241
Trinh, X.-T., Long, N.-V., Van Anh, L. T., Nga, P. T., Giang, N. N., Chien, P. N., Nam, S.-Y., & Heo, C.-Y. (2022). A comprehensive review of natural compounds for wound healing: Targeting bioactivity perspective. International Journal of Molecular Sciences, 23(17), 9573. https://doi.org/10.3390/ijms23179573van den Borne, S. W., van de Schans, V. A., Strzelecka, A. E., Vervoort-Peters, H. T., Lijnen, P. M., Cleutjens, J. P., Smits, J. F., Daemen, M. J., Janssen, B. J., & Blankesteijn, W. M. (2009). Mouse strain determines the outcome of wound healing after myocardial infarction. Cardiovascular Research, 84(2), 273-282. https://doi.org/10.1093/cvr/cvp207
Wei, T., & Dai, H. (2014). Quantification of GFP signals by fluorescent microscopy and flow cytometry. Methods in Molecular Biology , 1163, 23–31. https://doi.org/10.1007/978-1-4939-0799-1_3
Wolberg A. S. (2023). Fibrinogen and fibrin: synthesis, structure, and function in health and disease. Journal of thrombosis and haemostasis, 21(11), 3005–3015. https://doi.org/10.1016/j.jtha.2023.08.014