Gabe Anoos, Mercy Awosoga, and Anna Billones, Catholic Central High School, Lethbridge, Alberta, Canada
Amira Bach, Jovin Bach, Aysha Al Khater, Dana Omar, and Raafae Rasool, Chinook High School, Lethbridge, Alberta, Canada
Elyse Burgess, Nicole Burgess, Charlotte Fink, Saachee Josan, Nora Laird, Agrim Paudel, Jacqueline Sadr, and Maya Walton, Lethbridge Collegiate Institute, Lethbridge, Alberta, Canada
Ian Benkoczi, Mazen Chemissany, Avinash Chowdhury, Amryn Mutti, Selina Sun, Jerry Wang, Kathleen Wu, and Bobby Zhao, Winston Churchill High School, Lethbridge, Alberta, Canada
Reviewed on 3 May 2025; Accepted on 9 June 2025; Published on 27 October 2025
With help from the 2025 BioTreks Production Team.
In Alberta, more than 14,000 farmers grow over 6.6 million acres of canola every year. Canola is Canada’s most valuable crop, as it contributes over $29.9 billion annually to the economy. However, increasing challenges with climate change threaten the productivity of this sector. Conditions such as persistent drought have become a great concern to canola producers. Drought-resistant canola can be engineered through a collective approach combining plant-growth-promoting rhizobacteria and RNA interference (RNAi) to silence negative regulators of drought tolerance. A 2020 study using the model plant Arabidopsis thaliana showed that overexpression of bHLH61 impaired plant drought tolerance. However, the plants showed increased drought tolerance when this gene was mutated. There is a homologous gene found in canola, and we predict that by knocking down the expression of bHLH61 in canola, we can improve the drought tolerance of the crop. We will engineer Arthrobacter globiformis to express small-interfering RNA (siRNA) targeting the bHLH61 gene and incorporate the bacteria into the seed coating. The engineered bacteria can colonise the canola roots, delivering double-stranded RNA (dsRNA) to silence the bHLH61 gene while concurrently activating host stress response pathways to improve drought tolerance.
Keywords: Canola, drought, RNAi, siRNA, interference, knockdown, seed coating
Authors are listed in alphabetical order. Laura Keffer-Wilkes mentored the group. Please direct all correspondence to kefferwilkesl@uleth.ca.
Background
Southern Alberta’s agriculture sector is a crucial part of the Canadian and Alberta economy. It is characterized by expansive farmlands, diverse crop production, and advanced agricultural practices. The region includes over 10 million acres of farmland, representing more than 20% of Alberta’s agricultural land, and contributes over $2.6 billion annually in agricultural revenue (Stahl & Gething, 2024). Between 2021 and 2023, there has been a 24% growth observed in Alberta’s agricultural industry, making it even more crucial for the protection of this sector (Stahl & Gething, 2024). Increasing challenges with climate change threaten the productivity of Alberta’s agriculture sector. In recent years, conditions such as persistent drought, increasing pest pressures on crop production, winds, and lack of water have become of great concern to Alberta’s industry (Nickel & Williams, 2024). Historically, the area’s nutrient-rich soils have facilitated its development for livestock production, crop production, and agricultural processing, contributing $17.9 billion to Canada’s agrifood exports in 2023 (Calgary Economic Development, n.d.). This agricultural prominence is supported by a strong workforce and collaborative networks across sectors of research, technology, and industry, making Southern Alberta a critical driver of food security and economic stability in Canada.
In recent years, Southern Alberta has faced significant drought challenges exacerbated by consecutive dry years, below-average snowpack, and low reservoir levels. These below-average levels have placed Southern Alberta’s agricultural sector into Stage 4 of 5 in Alberta’s Water Shortage Management Plan (Government of Alberta, 2025). This stage is defined by water shortage in multiple water management areas, including reservoirs, lakes, and rivers, persistent water shortage over a lengthened period of time, and a significant number of agricultural users and households are severely impacted or unable to access water (Government of Alberta, 2023). The agriculture sector, heavily reliant on adequate irrigation and water supply, is now faced with soil erosion risks, limitations on water usage, and therefore water allocations (Stephenson, 2024).
Canola is one of Southern Alberta’s main agricultural crops. Canola production, cultivated across over 6.6 million acres by approximately 14,000 farmers, contributes $4 billion annually to Alberta’s agricultural revenue (Canola Statistics – Alberta Canola, n.d.). The crop’s prominence stems from its role as a high-value export commodity across Canada and on the global market. However, prolonged drought poses severe risks. Recent forecasts predict a 30-40% drop in Canadian canola production due to water scarcity, with southern Alberta particularly vulnerable given its semi-arid climate and lack of natural water bodies (Farmonaut, 2024). Drought conditions during early growth stages of the canola plants reduce seed yield and oil content, leading to extreme losses in revenue. Current research into drought-tolerant canola varieties is ongoing, but persistent drought conditions threaten the crop’s economic viability. Canola requires at least 400 mm of water to sustain growth throughout the entire life cycle, making insufficient water a key issue (Alberta Agriculture & Rural Development, 2011).
The current solutions are government-imposed water-sharing agreements and various agricultural technologies. In April 2024, major water users in the South Saskatchewan River Basin signed a large water-sharing agreement with Alberta to manage drought risk (Alberta Law Review, 2024). These agreements are intended to effectively use limited water supplies and reduce potential impacts on communities. However, this means that territories and communities are fully dependent on other land areas, which can be helpful in the short term but can be considered unreliable for several reasons. For example, because there is a limited supply of water, depending on other provinces can serve as only a temporary fix. Furthermore, the increasingly unpredictable weather patterns due to climate change can cause the current solutions to be outdated and ineffective.
The physiological responses in canola (Brassica napus) due to drought conditions affect many processes, including photosynthesis, stomatal conductance, transpiration, protein synthesis, and metabolite accumulation (Zhu et al., 2016). All these pathways either directly or indirectly affect seed yield and quality. The bHLH family of proteins plays an important role in plant stress response, but their exact molecular mechanism is still largely unknown. A 2020 study using the model plant Arabidopsis thaliana showed that overexpression of bHLH61 impaired plant drought tolerance, but when this gene was mutated, the plants showed increased drought tolerance. There is a homologous gene found in canola that codes for a hypothetical transcription factor and has been shown to be involved in cold tolerance previously (Wang et al., 2022). We predict that by knocking down the expression of bHLH61 in canola, we can improve the drought tolerance of the crop (Wang et al., 2021).
Systems Level
Our project proposes to knock down the expression of bHLH61 in canola using bacteria-mediated RNAi (Figure 1). Our engineered bacteria will be incorporated into the canola seed coating, then incorporated into the plant’s rhizosphere, where they will produce double-stranded hairpin RNA that targets the bHLH61 gene in canola, thus increasing drought tolerance.
| Figure 1. RhizoRetention proposes to engineer naturally symbiotic bacteria to deliver dsRNA to canola plants through the root system that will knock down expression of bHLH61 to increase drought tolerance. |
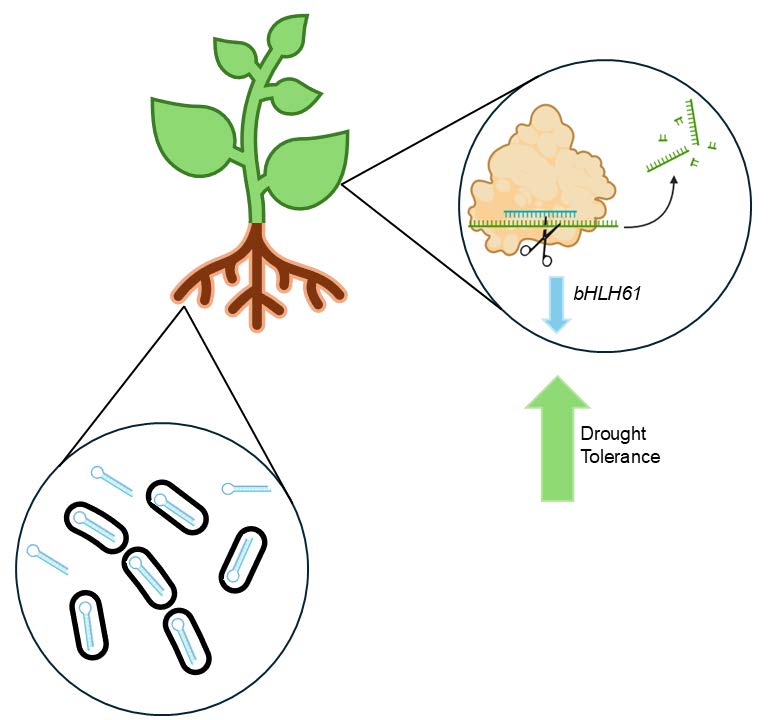
|
Device Level
Rhizosphere Microbes
Microorganisms will inhabit plant root environments and can form a symbiotic relationship with their host organism. Some microorganisms can help facilitate nutrient uptake, prevent colonization by pathogens, and help mitigate environmental stress factors’ impact (Lay et al., 2018). An analysis of canola root microbiomes in 2018 showed many rhizobacteria species, including Amycolatopsis, Serratia proteamaculans, Pedobacter, Arthrobacter, Stenotrophomonas, Fusarium merismoides, and Fusicolla (Lay et al., 2018). For our project, we aim to engineer and use Arthrobacter globiformis to express siRNA targeting canola stress response genes, and the bacteria will be incorporated as part of the seed coating.
Arthrobacter is a genus of Gram-positive soil bacteria that is often found as part of the indigenous bacterial biota of soil and rhizospheres. It is a native rhizobacteria in the canola root microbiome, which supports effective root colonization and long-term stability in the soil. Common microbiology and molecular biology techniques can be used to culture and transform Arthrobacter sp. with plasmids of interest. It is a biosafety level one organism and poses no threat to human health or the environment. However, in order for Arthrobacter to produce the siRNA, the RNase III gene will have to be deleted from the genome.
Seed Coatings
Seed coating is a process that consists of covering seeds with low amounts of exogenous materials, including insecticides, fungicides, nutrients, and biostimulants. Incorporating plant beneficial microbes into seed coatings can lead to increased germination, higher yields, and fewer fertilization needs. The engineered Arthrobacter bacteria would be included in the canola seed coating to allow for a cost-effective delivery system.
For effective water retention in plants, biopolymer-based hydrogel seed coatings are the best option, especially those that mimic natural seed mucilage. These are programmable coatings that have the ability to initiate rhizobacteria after sowing to form symbiotic water-stress-tolerant root nodules, especially in semi-arid environments (Zvinavashe et al., 2021). Sodium alginate and pectin hydrogels also show satisfactory performance, with excellent water holding capacity and maintaining favorable microbial activity for over one month. When fortified with NPK and micronutrients, these coatings significantly promote root and sprout development (Skrzypczak et al., 2021). While starch-based hydrogels help in initial growth under moderate water stress, their influence is lost once the plant has formed roots. Overall, hydrogel-based coatings can be tailored with beneficial microbes and can improve plant performance and water use.
RNAi
RNAi is a naturally occurring immune response against viral infections and can occur through multiple pathways (Baulcombe, 2004). Our team plans to utilize the siRNA pathway to target a gene within canola that is related to the expression of stress response proteins, specifically bHLH61. The siRNA is generated through transcription from a plasmid as a double-stranded hairpin precursor (Figure 2). It is then processed into the active siRNA sequence by Dicer. The siRNA is then integrated into the RNA-induced silencing complex (RISC), which can then scan mRNA to find the correct corresponding target sequence. Upon base pairing, the messenger RNA (mRNA) is degraded, leading to a loss of protein expression in the target plant (Svoboda, 2020).
| Figure 2. RNAi pathway using small interfering RNA against a target plant gene. The siRNA precursor hairpin is transcribed from the plasmid and then processed by Dicer (blue). The mature siRNA is then incorporated into RISC (tan) to degrade the target mRNA sequence. |
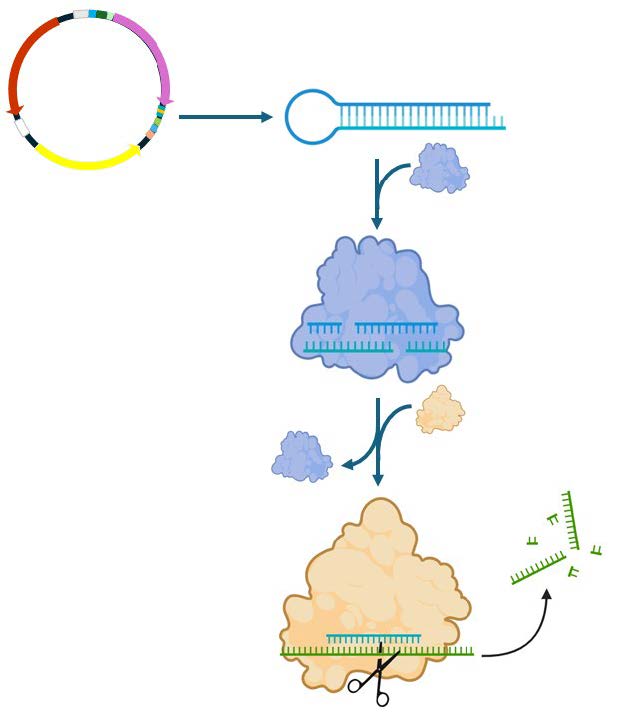
|
bHLH Super Gene Family
The basic helix-loop-helix (bHLH) gene family codes for the largest group of transcription factors in plants, meaning they help turn genes on and off in plants, but their functions are poorly characterized. In canola (Brassica napus), more than 600 potential bHLHs have been identified and categorized into 35 subfamilies (Ke et al., 2020). The bHLH gene family is highly conserved across plant species and is involved in growth and development, responses to light, drought, and pests, among others.
A 2020 study using the model plant Arabidopsis thaliana showed that overexpression of bHLH61 impaired plant drought tolerance, but when this gene was mutated, the plants showed increased drought tolerance (Wang et al., 2021). There is a homologous gene found in canola that codes for a hypothetical transcription factor (NIH, n.d.) and has previously been shown to be involved in cold tolerance in the related cultivar Brassica campestris (Wang et al., 2022).
Since transcription factors, like bHLH61, often have multiple roles, and knocking them down could lead to unintended consequences, transcriptomic profiling will be used to assess global gene expression changes and compared to the control samples. Additionally, phenotypic analyses under different stress conditions will be completed to hopefully catch any context-specific effects like cold sensitivity.
Parts Level
siRNA Design
The Arabidopsis gene bHLH61 is a potential target for RNA interference. The Arabidopsis locus tag for this gene is AT5G10570 and encodes for a putative myo-inositol hexakisphosphate kinase (NIH, n.d.). A standard nucleotide BLAST was completed using the Arabidopsis gene sequence against Brassica napus, and a predicted transcription factor bHLH61 (LOC106435320) gene was found to share 86% identity. For siRNA design, Gen siRNA Wizard (https://www.invivogen.com/sirnawizard/siRNA.php) was used to generate potential siRNA candidate sequences. These sequences were manually checked for any off-target sequence overlap using BLAST. Four candidate siRNA sequences were chosen to target the bHLH61 gene in canola (Table 1). Scramble sequences were made from the candidate sequences and will be used as negative controls to ensure that any observed effects are due to specific gene silencing and not caused by non-specific siRNA activity or the transfection process itself. We aim to use Arabidopsis as a proof-of-concept; therefore, siRNAs will be designed for both canola and Arabidopsis gene sequences. Additionally, for our proof-of-concept testing, we will create an siRNA that will target GFP mRNA in transgenic Arabidopsis.
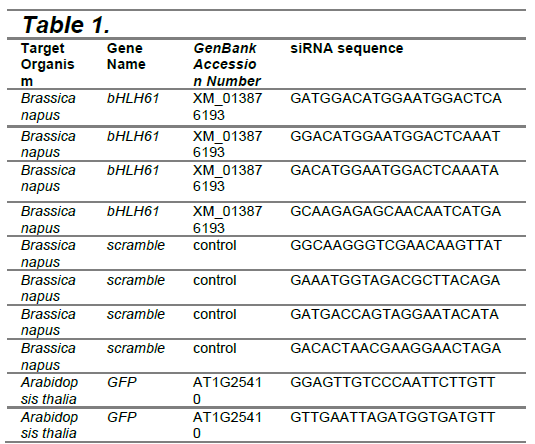
|
siRNA Production in Bacteria
We will base our plasmid design on the method used by Rueda-Silva (2023) and construct a novel plasmid using tsPurple, a chromoprotein, as a reporter gene to verify siRNA transcription (Figure 3). The new plasmid pPSLA will use the high-copy number backbone (pSB1C3), the T7 promoter (BBa_I712074) and terminator (BBa_K2246001), a strong ribosome binding site (RBS) (BBa_B0034), and the tsPurple chromoprotein coding region (BBa_K1033906). The different siRNA (sense, loop, antisense) sequences will immediately follow the tsPurple coding sequence, tying their transcription together. Any bacteria transformed with the plasmid must be deficient in RNase III to prevent degradation of the siRNA hairpin transcript.
| Figure 3. Plasmid pPSLA design overview. The plasmid contains a T7 promoter, RBS, tsPurple coding sequence, and the sense, loop, and antisense sequences for siRNA production. |
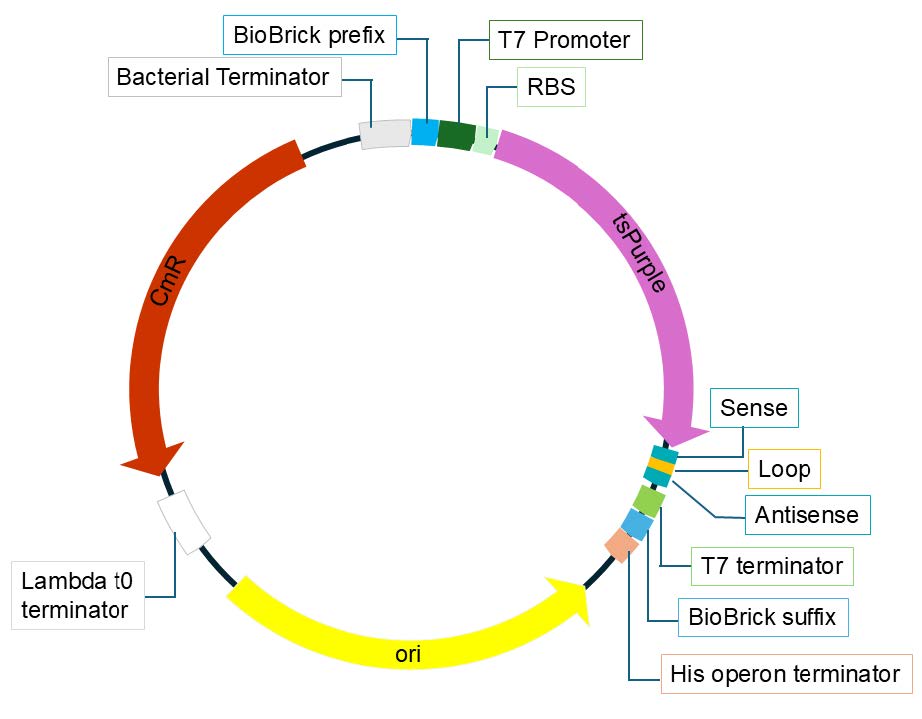
|
Safety
Three broad areas to be addressed by biosafety: containment of the genetically modified microorganism, environmental and public health, and regulation. The host organism, Arthrobacter globiformis strain ATCC 8010, is properly described by Environment and Climate Change Canada with no known negative effects on terrestrial/aquatic ecosystems or human communities and no in vitro pathogenicity or toxigenicity. Its occurrence in nature in soil and rhizospheres and a lack of growth capacity above 32°C limit growing potential in warm-blooded hosts or in non-target habitats (Environment and Climate Change Canada, 2018). In order to further inhibit horizontal gene transfer or survival, the engineered strain can possess auxotrophic deficiencies (i.e., artificial nutritional deficiency) or kill switches with inducibility, though the latter feature is not yet incorporated in the current design (Halvorsen et al., 2022)..
Risk to the environment is similar to the specificity of non-target organisms (NTOs) and RNAi activity, even though A. Globiformis per se has low risk; off-target activity of the RNAi construct has to be considered carefully. Bioinformatic tools are employed to design hairpin RNA to specifically target bHLH61 and minimize off-target gene silencing. RNAi crop research emphasizes the necessity of evaluating NTO routes of exposure, i.e., ingestion of plant material or soil residues, and associated RNAi induction in non-target organisms. For instance, dsRNA soil persistence and potential soil invertebrate (e.g., Folsomia candida) or aquatic uptake are questionable since the unjustified silencing of genes would disrupt critical biological processes. (Roberts et al., 2015). The seed-coating delivery system proposed, however, confines bacterial colonization to the rhizosphere and reduces enormous environmental dispersal. High-risk assessment by Canadian (CFIA), EU (Directive 2001/18), and International (Cartagena Protocol) regulatory agencies is required for GMMs and RNAi technologies.
The research will be under CFIA’s Seeds Regulations for seed coatings, such as information regarding bacterial viability, gene transfer efficiency, and RNAi stability. EU guidance regarding dsRNA-based plant protection products relies on sequence specificity, exposure pathways, and NTO testing, whereas the U.S. EPA regulates RNAi products as biochemical pesticides, which are required to possess ecotoxicological data. Besides, food safety assessment of RNAi crops needs to confirm the absence of unintended changes in their metabolism, not such a major problem in this situation, as the changed bacteria will not be intended as human food. Collaborative work with regulators and transparent risk communication will be the key concerns to align policy and public interests (Roberts et al., 2015; Kleter, 2020).
Discussions
One application of RNAi has been in agricultural pest and disease control as a more versatile and environmentally-friendly alternative than traditional methods, such as crop breeding and chemical pesticides, which may turn out to be slow and inefficient (Goodfellow et al., 2019). A genetic engineering approach using RNAi enables specific adjustments to genomes without widespread change (Vélez Arango et al., 2024). RNAi has been used in a multitude of ways to combat various viruses, parasites, fungi, and insects. For example, RNAi has been shown to form components of fungal pathogenesis and defense systems within the plant, combating fungal infections. (Goodfellow et al., 2019) There has been extensive research on the use of RNAi for improving crop stress tolerance, yield, and disease resistance. In regard to abiotic stressors, such as drought in particular, RNAi has been used in O. sativa (rice) to downregulate the RACK1 gene (receptor for activated C-kinase 1), which resulted in increased drought stress tolerance (Rajput et al., 2021). Similarly, disrupting SQS (farnesyltransferase/squalene synthase) in O. sativa via RNAi improved drought tolerance as well (Rajput et al., 2021). Having such points of reference is essential in development.
Despite the advantages of using RNAi, there are certain challenges that must be addressed for effective usage, particularly regarding environmental conditions. Soil pH and moisture levels directly influence nutrient uptake. (O’Kennedy, 2022) Deviations from optimal pH, which lie in the range of 6.0 to 7.5 for most crops and soil microbes, can hinder nutrient uptake. For instance, a pH level outside of this range may impact the availability of essential nutrients such as nitrogen, phosphorus, and potassium. (O’Kennedy, 2022). Additionally, either excessive or insufficient soil moisture levels pose challenges to microbial survival (Scheberl et al., 2019). Both soil pH and moisture levels should be taken into account when implementing RNAi within agricultural settings, as they affect the effectiveness of rhizobacteria and RNAi. Regarding traditional microbial seed coatings, they may face challenges such as poor adhesion and reduced microbial survival rates during storage. (Gong et al., 2023). Furthermore, many conditions, such as seed exudates and environmental conditions, can adversely affect the coating of a seed when sown. (Callaghan et al., 2012). It should also be considered that while seed coating techniques can introduce advantages to plants during growth, they also introduce additional costs and further complicate agricultural practices. The production of these coatings requires specialized equipment, which has the potential to increase overall costs. (Gong et al., 2023). Furthermore, laboratory or greenhouse conditions, under which we envision researching, could vastly differ from real-world agricultural settings. Thus, multiple environmental factors should be considered when looking into the viability of RhizoRetention.
Next Steps
The next steps include contacting Dr. Elizabeth Shultz of the Department of Biological Sciences at the University of Lethbridge. Her research focuses on understanding the genome of Arabidopsis, and she can provide guidance on using Arabidopsis as our model organism. Our current plan is to produce siRNA that will target the GFP gene in transgenic Arabidopsis. By targeting the GFP gene, we will be able to visually confirm knockdown of the target gene using our siRNA. We will also target bHLH61 in Arabidopsis for knockdown. The siRNA itself will be produced using the Escherichia coli (E. coli) HT115 strain, as this strain is RNase III deficient and has previously been used to produce large amounts of siRNA in vivo (Rueda-Silva et al., 2023). The siRNA will be purified from the bacteria and applied directly to Arabidopsis, either by soaking the roots or spraying the plant.
Author Contributions
Video by M.A., and A.P. Background by N.B., E.B., M.C., S.S., and B.Z. Systems by A.C., I.B., and J.W. Device by A.B., A.M., S.J., and K.W. Parts by M.A. and A.B. Safety by J.B., R.R., J.S., and C.F. Discussion by M.W., G.A., and N.L. Next Steps by D.O., I.B., B.Z., and S.S.
Acknowledgements
We would like to thank the University of Lethbridge, SynBridge, the Synthetic Biology Makerspace, and the University of Lethbridge’s Chemistry and Biochemistry Department for their support and encouragement. We also greatly appreciate Mind Fuel and their support of our project through the Tech Futures Challenge.
References
Baulcombe, D. (2004, November 16). RNA silencing in plants. Nature, 431, 356-363.
Calgary Economic Development. (n.d.). Agribusiness: Calgary is an agriculture-based city. https://www.calgaryeconomicdevelopment.com/sectors/agribusiness/
Callaghan, M., Wright, D., Swaminathan, J., Young, S., & Wessman, P. (2012). Microbial inoculation of seed – issues and opportunities. Agronomy New Zealand, 42.
Canola Statistics – Alberta Canola. (n.d.). Alberta Canola Producers Commission. Retrieved April 25, 2025, from https://albertacanola.com/about/canola-statistics/
Darlington, M., Jurat-Fuentes, J. L., Kogel, K.-H., Rathore, K. S., Smagghe, G., & Whyard, S. (2024). RNA Interference in Agriculture: Methods, Applications, and Governance. CAST. https://doi.org/10.62300/IRNE9191
Environment and Climate Change Canada. (2018). Final screening assessment for Arthrobacter globiformis. https://www.canada.ca/en/environment-climate-change/services/evaluating-existing-substances/screening-assessment-arthrobacter-globiformis.html
Gong, M., He, J., Kong, M., Huo, Q., Jiang, Y., Song, J., Han, W., & Lv, G. (2023). A microencapsulation approach to design microbial seed coatings to boost wheat seed germination and seedling growth under salt stress. Frontiers Plant Science, 14.
Goodfellow, S., Zhang, D., Wang, M.-B., & Zhang, R. (2019). Bacterium-Mediated RNA Interference: Potential Application in Plant Protection. Plants, 8(12). https://www.mdpi.com/2223-7747/8/12/572
Government of Alberta. (2023, August). Water Shortage Management Stages. chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://open.alberta.ca/dataset/fa34c6e0-14e2-4eb4-b553-8c6e13bf122a/resource/488e9eef-0c58-4527-91c7-1be7680368f5/download/epa-water-shortage-management-stages-2023-08.pdf
Government of Alberta. (2025). Drought – Current Conditions. https://www.alberta.ca/drought-current-conditions
Halvorsen, T. M., Ricci, D. P., Park, D. M., Jiao, Y., & Yung, M. C. (2022). Comparison of kill switch toxins in plant-beneficial Pseudomonas fluorescens reveals drivers of lethality, stability, and escape. ACS Synthetic Biology, 11(11), 3785–3796. https://doi.org/10.1021/acssynbio.2c00386
Ke, Y.-Z., Wu, Y.-W., Zhou, H.-J., Chen, P., Wang, M.-M., Liu, M.-M., Li, P.-F., Yang, J., Li, J.-N., & Du, H. (2020). Genome-wide survey of the bHLH super gene family in Brassica napus. BMC Plant Biology, 20. https://bmcplantbiol.biomedcentral.com/articles/10.1186/s12870-020-2315-8#:~:text=Conclusion,in%20future%20gene%20function%20research.
Kleter, G. A. (2020). Food safety assessment of crops engineered with RNA interference and other methods to modulate the expression of endogenous and plant pest genes. Pest Management Science. 10.1002/ps. 5957
Lay, C.-Y., Bell, T. H., Hamel, C., Harker, K. N., Mohr, R., Greer, C. W., Yergeau, E., & St-Arnaud, M. (2018, June). Canola Root–Associated Microbiomes in the Canadian Prairies. Frontiers Microbiology, 9. https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2018.01188/full#B73
Nickel, R., & Williams, N. (2024, March 18). Farmers, oil drillers in parched Alberta brace for water shortage. Reuters. https://www.reuters.com/business/environment/farmers-oil-drillers-parched-alberta-brace-water-shortage-2024-03-18/
NIH. (n.d.). Nucleotide BLAST. https://www.ncbi.nlm.nih.gov/nuccore/XM_013876193.3?report=fasta
NIH. (n.d.). https://www.ncbi.nlm.nih.gov/gene/830922
O’Kennedy, S. (2022). Soil pH and its impact on nutrient availability and crop growth. International Journal of Geography, Geology and Environment, 4(2), 236-238.
Rajput, M., Choudhary, K., Kumar, M., Vivekanand, V., Chawade, A., Ortiz, R., & Pareek, N. (2021). RNA Interference and CRISPR/Cas Gene Editing for Crop Improvement: Paradigm Shift towards Sustainable Agriculture. Plants, 10(9). https://www.mdpi.com/2223-7747/10/9/1914
Roberts, A., Devos, Y., Lemgo, G. N.Y., & Zhou, X. (2015). Biosafety research for non-target organism risk assessment of RNAi-based GE plants. Frontiers Plant Science, 6. https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2015.00958/full
Rueda-Silva, J. C., Gonzalez-Campos, L. I., Duran-Armenta, L. F., Karam-Coppola, A., Antonio-Perez, A., Ordonez-Rodriguez, J., Saucedo-Tavitas, N. E., Rico-Torres, V., Cruz-Cruz, C., Garcia-Huante, Y. G., Amador-Molina, J. C., Tapia-Ramirez, J. I., & Torres-Huerta, A. L. (2023). Novel bacterial plasmid produces small interfering RNAs (siRNAs) that induce effective gene silencing in the Asian citrus psyllid Diaphorina citri. Electronic Journal of Biotechnology, 64, 59-68. https://doi.org/10.1016/j.ejbt.2023.03.006
Scheberl, L., Scharenbroch, B. C., Werner, L. P., Prater, J. R., & Fite, K. L. (2019). Evaluation of soil pH and soil moisture with different field sensors: Case study urban soil. Urban Forestry & Urban Greening, 38, 267-279. https://doi.org/10.1016/j.ufug.2019.01.001
Skrzypczak, D., Jarzembowski, L., Izydorczyk, G., Mikula, K., Hoppe, V., Mielko, K. A., Pudelko-Malik, N., Mlynarz, P., Chojnacka, K., & Witek-Krowiak, A. (2021). Hydrogel Alginate Seed Coating as an Innovative Method for Delivering Nutrients at the Early Stages of Plant Growth. Polymers, 13. https://doi.org/10.3390/polym13234233
Stahl, H., & Gething, K. (2024). Economic Impact of Agriculture on Southeast Alberta (2024). C4A Centre for Innovation, 1(1). file:///C:/Users/kefferwilkesl/Downloads/Economic%20Impact%20of%20Agriculture%202024.pdf
Stephenson, A. (2024, January 22). Farmers in Alberta face a growing risk of soil erosion events as drought persists. The Canadian Press. https://globalnews.ca/news/10241453/alberta-farmers-soil-erosion-risk/
Svoboda, P. (2020). Key mechanistic principles and considerations concerning RNA interference. Frontiers in Plant Science, 11, 1237. https://doi.org/10.3389/fpls.2020.01237
Vélez Arango, A. M., Darlington, M., Jurat-Fuentes, J. L., Kogel, K.-H., Rathore, K. S., Smagghe, G., & Whyard, S. (2024). RNA Interference in Agriculture: Methods, Applications, and Governance. Council for Agricultural Science and Technology. https://doi.org/10.62300/IRNE9191
Wang, C., Zhang, M., Zhou, J., Gao, X., Zhu, S., Yuan, L., Hou, X., Liu, T., Chen, G., Tang, X., Shan, G., & Hou, J. (2022). Transcriptome analysis and differential gene expression profiling of wucai (Brassica campestris L.) in response to cold stress. BMC Genomics, 23. https://bmcgenomics.biomedcentral.com/articles/10.1186/s12864-022-08311-3
Wang, Y., Fang, Z., Yang, L., & Chan, Z. (2021). Transcriptional variation analysis of Arabidopsis ecotypes in response to drought and salt stresses dissects commonly regulated networks. Physiologia Plantarum, 172(1), 77-90. https://onlinelibrary.wiley.com/doi/10.1111/ppl.13295
Zvinavashe, A. T., Laurent, J., Mhada, M., Sun, H., Fouda, H. M. E., Kim, D., Mouhib, S., Kouisni, L., & Marelli, B. (2021, July). Programmable design of seed coating function induces water-stress tolerance in semi-arid regions. Nature Food, 2, 485-493. https://doi.org/10.1038/s43016-021-00315-8