Frances Dunn, Tee Jarukulvanich, Tanya Jay, Tasheen Kashem, Yeseo Kim, Nathaniel Lawrence, Aarav Phadnis, Charvi Talupuru, Annabel Volpe, Lexington High School, Lexington, Massachusetts, United States
Reviewed on 3 May 2025; Accepted on 9 June 2025; Published on 27 October 2025
With help from the 2025 BioTreks Production Team.
Vitamin A deficiency is caused by an insufficient intake of vitamin A in the diet. This deficiency is a widespread health issue that particularly affects developing countries. A possible solution is the genetic engineering of crops containing vitamin A precursors, such as β-carotene, a type of carotenoid found in various fruits and vegetables that can be converted into vitamin A by the human body. The stability of carotenoids ultimately determines how much can be used by the human body, and is considered a critical issue in genetically engineered crops. Using the model plant Nicotiana benthamiana for rapid gene expression in plant biology, we aim to increase the stability of β-carotene in the cytosol of the plant to improve the nutritional availability of the vitamin and combat the global deficiency. Our project aims to introduce lipid droplets into the cytosol of N. benthamiana leaves, while simultaneously expressing four key genes that are involved in the synthesis of carotenoids in the cytosol: crtB (bacterial phytoene synthase), crtI (phytoene desaturase), crtE (lycopene cyclase) , and a cytosolic version of hydroxymethylglutaryl-CoA reductase (tHMGR). As lipid droplets provide a protective environment that reduces exposure to oxidative conditions, we hypothesize that their incorporation in the biofortified N. benthamiana leaves will improve the stability of β-carotenoids in the cytosol, leading to greater carotene availability and more efficient conversion to vitamin A upon human consumption.
Keywords: Carotenoids, Biofortification, Vitamin A, Vitamin A Deficiency (VAD), Nicotiana benthamiana
Authors are listed in alphabetical order. Reginald Hobbs mentored the group. Please direct all correspondence to rhobbs@lexingtonma.org.


Background
Vitamin A is a group of essential, fat-soluble micronutrients, including retinol, retinal, retinoic acid, and various provitamin A carotenoids, such as β-carotene, which is enzymatically cleaved and converted into retinol in the intestines and liver. These compounds are critical for a variety of biological processes, including vision (particularly in low-light conditions), immune system function, reproduction, and cellular communication. Retinoic acid, the active form of vitamin A, plays a key role in gene expression, influencing the growth and development of organs, epithelial integrity, and immune defense. Retinal is indispensable for the synthesis of rhodopsin, a pigment found in the retina that enables night vision. Given its broad physiological roles, vitamin A is vital for maintaining human health.(Goodman et al., 1966). Vitamin A is an essential nutrient, meaning it cannot be synthesized by the human body, and must instead be obtained from external dietary sources. These include preformed vitamin A from animal-derived foods like milk, eggs, and fish oils, and provitamin A carotenoids (particularly β-carotene) found in plant-based sources like carrots, cantaloupe, mango, and leafy greens (Akhtar et al., 2013). However, in many developing nations, access to vitamin A-rich foods is severely limited due to food insecurity, poverty, limited agricultural diversity, and poor infrastructure (Morelli et al., 2024). As a result, vitamin A deficiency (VAD) remains a critical public health concern, especially in sub-Saharan Africa and South Asia. According to the World Health Organization, VAD affects an estimated 190 million preschool-age children and 19 million pregnant women globally. The deficiency is most prevalent in children under the age of five, where it contributes to approximately 670,000 deaths annually due to its role in compromising immune responses to infectious diseases such as measles and diarrhea (Akhtar et al., 2013).
Figure 1 shows the prevalence of vitamin A deficiency in global populations, with the highest rates occurring in resource-poor regions. VAD is particularly dangerous for young children and pregnant women due to their elevated nutritional requirements. In children, VAD is the leading cause of preventable blindness and increases the risk of severe infections and death. Symptoms of VAD begin with night blindness and progress to xerophthalmia, a spectrum of eye disorders that includes conjunctival and corneal dryness, Bitot’s spots, and eventually, irreversible blindness from corneal ulceration and keratomalacia. In pregnant women, VAD can cause night blindness and increase maternal mortality due to complications during childbirth.(Akhtar et al., 2013).
| Figure 1. Map of vitamin A deficiency distribution worldwide (Dubock, 2024). |
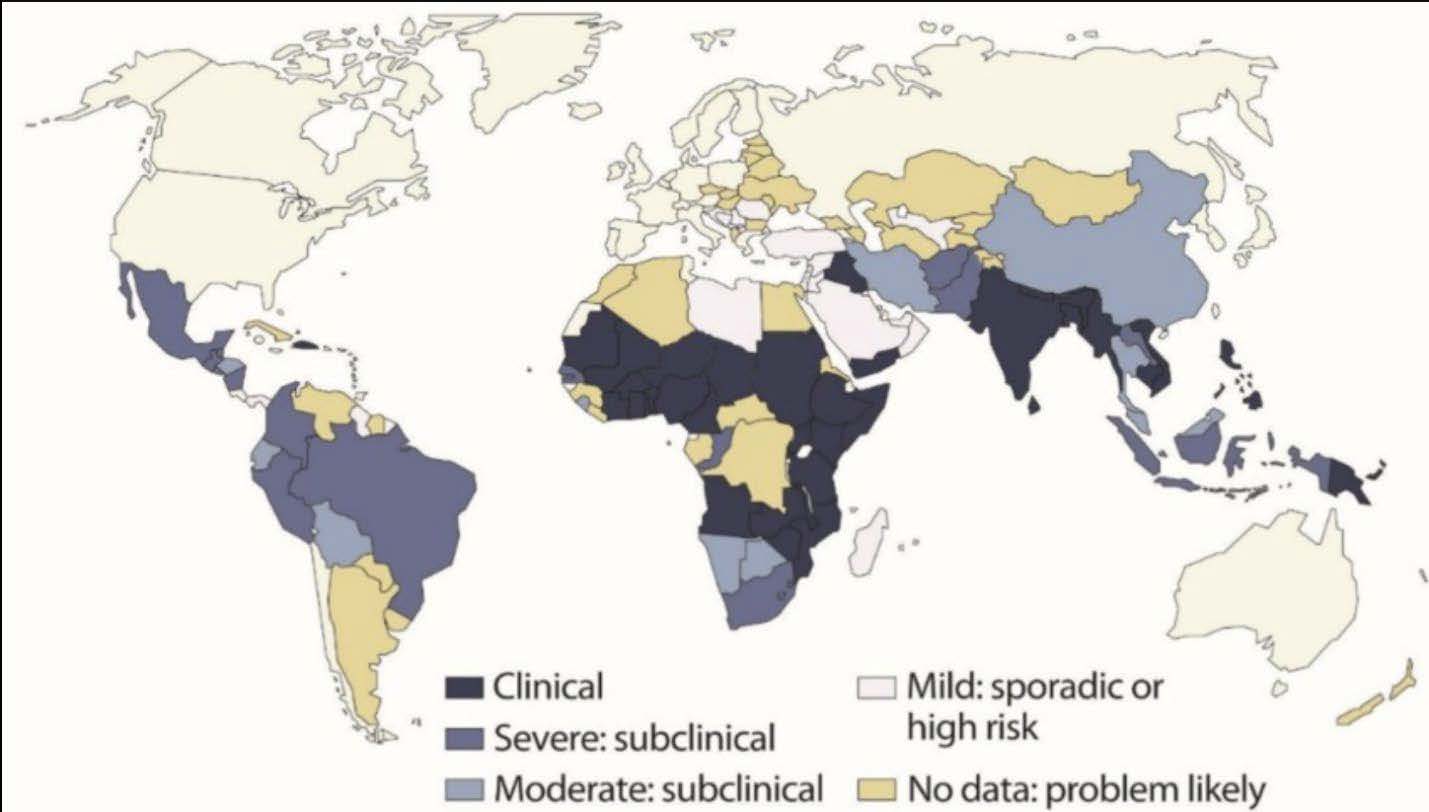
|
Efforts to combat VAD have included vitamin A supplementation, fortification programs, and dietary diversification. However, these strategies face several limitations in developing countries. Supplementation programs are often logistically challenging and expensive to maintain. Fortification is dependent on the availability of centralized food processing systems, which are frequently lacking in rural areas. Lastly, dietary diversification is constrained by local agricultural practices, cultural preferences, and seasonal availability of vitamin A-rich foods.
To address these limitations, recent biotechnological advances have turned toward biofortification, the genetic enhancement of staple crops to increase their micronutrient content. Golden Rice, a genetically modified rice variety enriched with β-carotene, was among the first successful examples of this approach (Potrykus, 2001). Our team seeks to adapt and expand this strategy using Nicotiana benthamiana as a model system to enhance both the stability and yield of β-carotene.
Specifically, our goal is to enhance β-carotene stability by introducing lipid droplets in the cytosol of N. benthamiana cells, creating a more favorable environment for carotenoid accumulation. β-carotene is a type of carotenoid pigment present in several fruits and vegetables. Once ingested, β-carotene is absorbed by the intestinal mucosa and cleaved by the enzyme beta-carotene 15,15’-monooxygenase to yield retinal, which is then reduced to retinol or oxidized to retinoic acid depending on the body’s needs. When expressed in the model plant N. benthamiana using four genes – crtB, crtB, crtE, and tHMGR, the plant shows increased β-carotene accumulation (Morelli et al., 2024). The genes crtB, crtB and crtE encode enzymes responsible for the biosynthesis of carotenoids, producing β-carotene from precursor molecules. Additionally, tHMGR is included to boost the supply of isopentenyl pyrophosphate (IPP), which is a necessary component of the biosynthesis production pathway (Cunningham et al., 1993; Enfissi et al., 2005). Despite increased synthesis, a major challenge remains: the intracellular instability of β-carotene. β-carotene is chemically unstable and highly prone to degradation when exposed to heat or oxidative stress, such as during storage or cooking, particularly in hot and humid environments. To address this, we propose a novel strategy to stabilize β-carotene by incorporating lipid droplets into the cytosol via agroinfiltration. Lipid droplets are intracellular organelles composed of a neutral lipid core surrounded by a phospholipid monolayer and associated proteins. These structures naturally serve as storage compartments for hydrophobic molecules, including carotenoids, and help protect them from degradation (Yan et al., 2024).
Unlike preformed vitamin A (retinol), which can be toxic in high doses, due to its fat-soluble nature and potential for accumulation in the liver, β-carotene, the key nutrient targeted in this design, is generally regarded as safe. This is because the body regulates its conversion to vitamin A based on physiological need, reducing the risk of hypervitaminosis A (Goodman et al., 1966; Shang et al., 2021).
The successful implementation of this strategy would provide a sustainable, cost-effective, and culturally appropriate method to increase vitamin A intake in vulnerable populations. Unlike synthetic supplements or fortified foods, biofortified plants can be propagated and harvested locally, empowering communities to take ownership of their nutritional needs. By increasing both the production and the resilience of β-carotene in plants, our approach offers a long-term solution to a persistent global health crisis.
Systems Level
To address global vitamin A deficiency through plant biofortification, our team has designed a synthetic biological system that operates within the cells of N. benthamiana leaves. The goal is to enhance both the accumulation and bioaccessibility of provitamin A carotenoids, particularly β-carotene. This system integrates genetic and metabolic engineering by introducing four key genes, crtB, crtL, crtE, and tHMGR, into the cytosol of N. benthamiana leaf cells. These genes drive the production of β-carotene through the carotenoid biosynthesis pathway (Sandmann, Misawa, & Bock, 1991; Cunningham et al., 1993; Enfissi et al., 2005).
To protect the synthesized β-carotene from oxidative degradation, we have also incorporated the co-expression of cytosolic lipid droplets that act as stabilizing carriers for the synthesized carotenoids. These lipid droplets function as storage compartments that are induced by agroinfiltration to sequester and stabilize the β-carotene molecules (Sadre et al., 2019; de Abreu Martins et al., 2021). Agroinfiltration is a transient gene expression technique in which Agrobacterium tumefaciens carrying the genetic constructs are infiltrated into plant tissues such as plant leaves, enabling the expression of multiple genes transiently (Chen et al., 2013; Su et al., 2023).
Figure 2. The 4 genes crtB, crtL, crtE, and tHMGR are introduced into N. Benthamiana leaves via agroinfiltration along with cytosolic lipid droplets and the RUBY construct for detection.
In addition, the central biosynthetic module, compatible with crtB, crtL, crtE, and tHMGR, is improved by the addition of extra genetic elements that trigger the formation of lipid droplets. Simply put, we co-express genes that encode proteins associated with lipid droplets, such as diacylglycerol acyltransferase (DGATs) and Oleosin proteins, along with the four carotenoid biosynthetic genes. These extra genes are organized into their own separate expression cassettes within the general construction and are delivered together through agroinfiltration. This design ensures that lipid droplets are made at the same time as β-carotene is produced, creating natural storage compartments that not only help stabilize carotenoids, but also protect them from oxidative damage, improving their bioaccessibility when consumed (de Abreu Martins et al., 2021; Kaur et al., 2024).
As a whole, the system operates by boosting the production of β-carotene via engineered metabolic pathways while simultaneously increasing its stability and thus its availability upon digestion. This 2-part approach not only raises the total carotenoid content within the plant but also enhances its bioavailability, making it more effective in providing a source of vitamin A (Morelli et al., 2024).
Device Level
The central component of this research is engineering a dual-functioning system within the N. benthamiana plant using the biosynthesis of β-carotene in the cytosol and the induction of cytosolic lipid droplets. N. benthamiana is an ideal model organism for this system on account of its proven susceptibility to Agrobacterium-mediated gene transformation and low peptidase profile, which allows for it to be rapidly transformed through transient gene expression and supports high levels of protein production within a short period (Goodin et al., 2008).
Our device-level implementation involves a step-by-step approach: i) the insertion of genes for β-carotene biosynthesis into the cytosol, achieved by agroinfiltration using Agrobacterium tumefaciens carrying plasmid-based expression vectors (these plasmids allows transient expression of crtB, crtl, crtE, and THMGR genes into the cytoplasm without integration into the genome (He et al., 2020; Morelli et al., 2024)), ii) addition of lipid droplets to enhance carotenoid stability, iii) using genes encoding diacylglycerol acyltransferase (DGATs) and Oleosin proteins, which promote the formation and stabilization of lipid droplets capable of sequestering β-carotene (Sadre et al., 2019; de Abreu Martins et al., 2021); and iv) the integration of a RUBY reporter system to visually confirm successful transformation. Drawing on techniques from Morelli et al. (2024) and Su et al. (2023), and utilizing transformation methods established by He et al. (2020), this design serves as a basis for a transposable biofortification platform.
By engineering a cytosolic environment optimized for carotenoid synthesis and storage, we aim to create a plant-based system for nutritional enhancement.
Parts Level
To achieve biosynthesis of β-carotene in the cytosol, a coordinated expression of four key genes, which are optimal for carotenoid biosynthesis, is employed. The bacterial gene crtB encodes phytoene synthase, an enzyme that catalyzes the conversion of pre-phytoene diphosphate to phytoene, the first step of carotenoid biosynthesis (Sandmann et al., 1991). Furthermore, the bacterial gene crtL encodes a desaturase enzyme that catalyzes the conversion of phytoene to lycopene, a key intermediate in the biosynthesis process (Armstrong et al., 1989). Lycopene cyclase, the enzyme encoded by the bacterial gene crtE, catalyzes the cyclization of lycopene into β-carotene by adding β-ionic rings to both ends of the molecule, a critical enzymatic step incorporated in our genetic design to enable sufficient β-carotene synthesis (Cunningham et al., 1993). To enhance the availability of cytosolic precursors for carotenoid biosynthesis, the pathway includes the addition of hydroxymethylglutaryl-CoA Reductase (tHMGR), which boosts levels of isopentenyl pyrophosphate (IPP), a key metabolic precursor (Enfissi et al., 2005).
Alongside reinforcing the β-carotene production in the cytosol, implementing a protective storage environment could be accomplished through introducing lipid droplets within the leaf cytosol, which results in gene expression known to promote lipid droplet formation. The study by Morelli et al. (2024) demonstrates a successful strategy of combining engineered biosynthesis and storage pathways to boost pro-vitamin A content in leaves, highlighting the possibility of mimicking similar storage and stability mechanisms for carotenoid accumulation. The potential for lipid droplets to stabilize lipophilic compounds is further supported by research that depicts how lipid nanoparticles protect β-carotene from degradation (de Abreu Martins et al., 2021). Drawing directly from the work of Su et al. (2023), which provides a comprehensive overview of engineering plant lipid droplets, our research explores the use of similar genetic constructs to induce lipid droplet formation in the cytosol. These constructs may comprise genes encoding key structural proteins of lipid droplets , such as oleosins, and enzymes involved in triacylglycerol synthesis, such as specific diacylglycerol acyltransferases (DGATs), potentially utilizing the same or highly homologous sequences identified and characterized in their study. Our hypothesis is that the lipid droplets will shield the carotenoids from the aqueous cytosolic environment and potential oxidative damage, which is a crucial factor considering the well-known instability of β-carotene (de Abreu Martins et al., 2021).
All genetic constructs – those for β-carotene biosynthesis (crtB, crtl, crtY, tHMGR) and lipid droplet formation (OLEOSIN, DGATs) are introduced in the N.benthamiana leaves through Agrobacteroim-mediated transient expression (Su et al., 2023). This strategy allows for the simultaneous expression of multiple genes, proving to be particularly efficient for metabolic engineering projects (Chen et al., 2013; Su et al., 2023). The process first involves cloning each target gene (crtB, crtl, crtY, tHMGR, OLEOSIN, and DGAT) into plasmids compatible with the Agrobacterium. The plasmids are then transformed into Agrobacterium tumefaciens using electroporation or freeze thaw methods, routinely used for Agrobacterium transformation (Rahman et al., 2024). Once each strain has reached consistent gene expression levels following a growth stage, equal volumes of the strains are combined and injected into N. benthamiana leaves using a needleless syringe (Goodin et al., 2008; Golubova et al., 2024). It is estimated the induced genes will begin to be expressed 2 to 5 days after infiltration (Chen et al., 2024; He et al., 2020).
To facilitate the visualization of results, a RUBY reporter system is employed. The RUBY construct is a synthetic gene that is commonly used in experiments in the field of plant biotechnology. The system monitors transformations within the plant due to gene expression, by encoding the CYP76AD1 gene (responsible for betalain biosynthesis), DODA (dioxygenase involved in betalain synthesis), and a glucosyltransferase, which allows for the conversion of tyrosinase into detectable red betalain pigments (He et al., 2020). The utility of the RUBY reporter for visualization of the transgenic expression has been demonstrated in soybean gene editing by Chen et al.(2024).This red pigmentation offers a significant advantage as a non-invasive marker, unlike reporters like GUS or luciferase, leaving transgenic analysis to be visual and non-destructive (He et al., 2020). Utilizing the reporter allows us to correlate the presence of the red RUBY reporter signal with the efficiency of the β-carotene biosynthesis pathway and the formation of the engineered lipid droplets.
Safety
While this study focuses on improving the nutritional content of plants through provitamin A biofortification, safety remains a central consideration throughout the design, testing, and future deployment phases.
Although β-carotene is mainly safe, a trial run by the ATBC indicated that high-dose β-carotene supplementation (20 mg per day), with or without 50 mg per day of vitamin E, for 5-8 years raised the risk of lung cancer mortality among male smokers. The trial similarly found that combining high doses of β-carotene with 7,500 µg RAE of retinyl palmitate for 4-8 years in current and former smokers, as well as men exposed to asbestos, also increased the risk of getting and dying from lung cancer. However, these findings relate to purified, high-dose supplements rather than dietary or plant-based sources like the ones we hope to implement (Shang et al., 2021).
Our design uses N. benthamiana, a non-food model plant, to minimize ecological risk and prevent unintended consumption during early testing (Golubova et al., 2024). To prevent the environmental spread of genetically modified plant material, we plan to implement some biological containment strategies. These include the physical isolation from pollinators and from open-air environments during the growth and testing phase. We also use transient growth systems (Agrobacterium mediated infiltration) without stable genome integration, to ensure that the transformation is solely temporary and does not pass to future plant generations.
Nevertheless, as our research progresses toward edible crops, further studies and regulatory evaluations will be required to establish safe consumption thresholds and assess population-specific risks, particularly for vulnerable groups such as pregnant women or individuals with existing smoking-related health concerns.
Discussions
This investigation highlights the potential of biofortification and genetic engineering as promising tools in the global fight against vitamin A deficiency. Our work explores a novel approach to improving the nutritional value of plants through the simultaneous expression of genes involved in β-carotene biosynthesis in N. benthamiana. The anticipated outcome, the successful accumulation and stabilization of β-carotenoids in the cytosol, is supported by findings from prior research, such as Morelli et al. (2024), which emphasizes the importance of minimizing oxidative degradation to retain carotenoid content.
One of the major advantages of our system is its use of horizontal gene expression through agroinfiltration, which offers a rapid and flexible method for testing gene function. This allows for fast iteration cycles during the development process and is especially effective due to N. benthamiana’s susceptibility to Agrobacterium-mediated transformation (Goodin et al., 2008; Golubova et al., 2024). Additionally, the cytosolic localization of β-carotene offers a potentially more accessible and digestible form for eventual human consumption (Morelli et al., 2024).
Despite these strengths, the system is not without its challenges. A primary limitation lies in the need for repeated agroinfiltration, which can lead to inconsistent gene expression and is not easily scalable for agricultural deployment. Environmental factors such as temperature, humidity, and plant age can also influence transformation efficiency and expression levels, making reproducibility more difficult in field conditions (He et al., 2020; Su et al., 2023).
Looking forward, future work should focus on overcoming these limitations by exploring stable genetic transformation methods, such as Agrobacterium-mediated transformation or CRISPR-based gene integration, improving gene expression consistency through the use of inducible promoters, and adapting the system for long-term use in edible crops by testing in model food plants like tomato or rice (Zhou et al. 2020; Zhang et al., 2018; Rahman et al., 2024).
Expanding the study beyond the laboratory will be essential for understanding how this technology performs under real-world agricultural conditions and in diverse environmental settings. Ultimately, this design sets the stage for broader applications in plant-based nutrition and opens the door for further refinement of genetically engineered systems aimed at improving global health.
Next Steps
To validate our design, we will implement our experiment design in N. benthamiana leaves to assess the effects of cytosolic lipid droplets on carotenoid stability and bioaccessibility. The experiment begins with a plasmid containing four key genes in carotenoid biosynthesis, crtB, crtI, crtE, and tHMGR, which have been shown to increase β-carotene accumulation in the cytosol (Sandmann et al., 1991; Armstrong et al., 1989; Cunningham et al., 1993; Enfissi et al., 2005). These constructs, along with a gene known to induce lipid droplet formation in plant cells, will be introduced into Agrobacterium tumefaciens cultures, which will serve as the delivery vehicle for genetic transformation via agroinfiltration (Golubova et al., 2024).
N. benthamiana plants will then be grown under controlled conditions and co-infiltrated with the agrobacterium cultures using a needleless syringe (Goodin et al., 2008). Leaves will be separated into three experimental groups: leaves receiving both the carotenoid and lipid droplet constructs, leaves receiving only the carotenoid constructs (as a positive control group as demonstrated in Morelli et al., 2024), and infiltrated leaves (as a negative control group as demonstrated in Morelli et al., 2024). After transformation, plants will be maintained under a two-step light treatment, initially high light intensity (W500), followed by moderate light exposure (W50), to promote optimal gene expression and carotenoid accumulation (Morelli et al., 2024).
To test the experimental outcomes, leaf discs will be harvested weekly using an Eppendorf tube to apply pressure and collect consistent samples. These discs will then be dried and analyzed for carotenoid content via high-performance liquid chromatography, a standard for quantifying pigments (Su et al., 2023). For lipid droplet analysis, droplets will be isolated from the leaf tissue and visualized using centrifugation, followed by lipid extraction by thin-layer chromatography and mass spectrometry to confirm successful formation and assess droplet composition (Sadre et al., 2019). To evaluate the bioaccessibility of β-carotene, in vitro digestion assays will be conducted, comparing samples across all three treatment groups. The assays will be performed using the gastrointestinal digestion model (Minekus et al., 2014).
We anticipate that the co-expression of lipid droplets will enhance both the accumulation and bioaccessibility of β-carotene in plant tissues, as the droplets are expected to encapsulate and thus protect the carotenoids from oxidative degradation (Sadre et al., 2019; de Abreu Martins et al., 2021). If successful, this experiment could offer a novel strategy for improving the nutritional profile of leafy vegetables and set the groundwork for future applications in food crops such as plantain, which could serve as a sustainable solution for addressing nutrient deficiencies in under-resourced regions.
Author Contributions
All authors made contributions to the conceptualization, design, and execution of this project. Each author was involved in writing the various sections of the manuscript.
Acknowledgements
We would like to thank AoPS Lexington for sponsoring our participation in the BioBuilder Club program and making this project possible for our team. We are especially grateful to Dr. Tianhu Sun for his valuable guidance in developing our project idea and helping us design our experiment. We also deeply appreciate Mr. Hobbs for his continuous support throughout the process, assisting us in our project, keeping us on track, and teaching us the foundational concepts in biology and scientific communication we needed to succeed. We would also like to thank the BioBuilder Foundation for their support and assistance in navigating the design process and for connecting us with our mentor.
References
Akhtar, S., Ahmed, A., Randhawa, M. A., Atukorala, S., Arlappa, N., Ismail, T., & Ali, Z. (2013). Prevalence of vitamin A deficiency in South Asia: Causes, outcomes, and possible remedies. Journal of Health, Population, and Nutrition, 31(4), 413–423. https://doi.org/10.3329/jhpn.v31i4.19975
Armstrong, G. A., Alberti, M., Leach, F., & Hearst, J. E. (1989). Nucleotide sequence, organization, and nature of the protein products of the carotenoid biosynthesis gene cluster of Rhodobacter capsulatus. Molecular and General Genetics, 216(2–3), 254–268. https://doi.org/10.1007/bf00334364
Cai, Y., Goodman, J. M., Pyc, M., Mullen, R. T., Dyer, J. M., & Chapman, K. D. (2015). Arabidopsis SEIPIN Proteins Modulate Triacylglycerol Accumulation and Influence Lipid Droplet Proliferation, The Plant Cell, 27(9), 2616–2636. https://doi.org/10.1105/tpc.15.00588
Chen, L., Cai, Y., Liu, X., Yao, W., Wu, S., & Hou, W. (2024). The RUBY reporter for visual selection in soybean genome editing. aBIOTECH, 5(2), 209–213. https://doi.org/10.1007/s42994-024-00148-6
Cunningham, F. X., Jr., Chamovitz, D., Misawa, N., Gantt, E., & Hirschberg, J. (1993). Cloning and functional expression in Escherichia coli of a cyanobacterial gene for lycopene cyclase, the enzyme that catalyzes the biosynthesis of β-carotene. FEBS Letters, 328(1–2), 130–138. https://doi.org/10.1016/0014-5793(93)80980-9
de Abreu Martins, H. H., Turmo-Ibarz, A., Hilsdorf Piccoli, R., Martín‐Belloso, O., & Salvia-Trujillo, L. (2021). Influence of lipid nanoparticle physical state on β-carotene stability kinetics under different environmental conditions. Food & Function, 12(2), 840–851. https://doi.org/10.1039/d0fo01980a
De Moura, F. F., Moursi, M., Donahue Angel, M., Angeles-Agdeppa, I., Atmarita, A., Gironella, G. M., Muslimatun, S., & Carriquiry, A. (2016). Biofortified β-carotene rice improves vitamin A intake and reduces the prevalence of inadequacy among women and young children in a simulated analysis in Bangladesh, Indonesia, and the Philippines. The American Journal of Clinical Nutrition, 104(3), 769–775. https://doi.org/10.3945/ajcn.115.129270
Díaz-Sánchez, V., Estrada, A. F., Trautmann, D., Limón, M. C., Al-Babili, S., & Avalos, J. (2011). Analysis of al-2 mutations in Neurospora. PLOS ONE, 6(7), e21948. https://doi.org/10.1371/journal.pone.0021948
Dubock, A. (2024). The Sustainable Development Goals are Unattainable without a Change of Nutritional Strategy. Medical Research Archives, 12(10). https://doi.org/10.18103/mra.v12i10.5818
Enfissi, E. M. A., Bramley, P. M., Fraser, P. D., & Valpuesta, V. (2005). Metabolite profiling of fruits of tomato lines differing in ploidy reveals marked changes in primary and secondary metabolism. Plant Physiology, 137(4), 1351–1362.
Golubova, D., Tansley, C., Su, H., & Patron, N. J. (2024). Engineering Nicotiana benthamiana as a platform for natural product biosynthesis. Current Opinion in Plant Biology, 81, 102611. https://doi.org/10.1016/j.pbi.2024.102611
Goodin, M. M., Zaitlin, D., Naidu, R. A., & Lommel, S. A. (2008). Nicotiana benthamiana: Its history and future as a model for plant-pathogen interactions. Molecular Plant-Microbe Interactions, 21(8), 1015–1026. https://doi.org/10.1094/MPMI-21-8-1015
Goodman, D. S., Huang, H. S., & Shiratori, T. (1966). Mechanism of the biosynthesis of vitamin A from beta-carotene. The Journal of Biological Chemistry, 241(9), 1929–1932. https://doi.org/10.1016/S0021-9258(18)96648-2
He, Y., Zhang, T., Sun, H., Zhan, H. & Zhao, Y. (2020). A reporter for noninvasively monitoring gene expression and plant transformation. Horticulture Research, 7, 152. https://doi.org/10.1038/s41438-020-00390-1
Jing, Y., Wang, J., Gao, H., Jiang, Y., Jiang, W., Jiang, M., Xin, F., & Zhang, W. (2023). Enhanced β-carotene production in Yarrowia lipolytica through metabolic and fermentation engineering. Journal of Industrial Microbiology & Biotechnology, 50(1), kuad009. https://doi.org/10.1093/jimb/kuad009
Kaur, M., Sinha, K., Eastmond, P. J. & Bhunia, R. K. Exploiting lipid droplet metabolic pathway to foster lipid production: oleosin in focus. Plant Cell Rep 44, 12 (2025). https://doi.org/10.1007/s00299-024-03390-w
Minekus, M., Alminger, M., Alvito, P., Ballance, S., Bohn, T., Bourlieu, C., Carrière, F., Boutrou, R., Corredig, M., Dupont, D., Dufour, C., Egger, L., Golding, M., Karakaya, S., Kirkhus, B., Le Feunteun, S., Lesmes, U., Macierzanka, A., Mackie, A., Marze, S., McClements, D. J., Ménard, O., Recio, I., Santos, C. N., Singh, R. P., Vegarud, G. E., Wickham, M. S. J., Weitschies, W. & Brodkorb, A. (2014). A standardised static in vitro digestion method suitable for food – an international consensus. Food & Function, 5(6), 1113–1124. https://doi.org/10.1039/c3fo60702j
Morelli, L., Perez-Colao, P., Reig-Lopez, D., Di, X., Llorente, B., & Rodriguez-Concepcion, M. (2024). Boosting pro-vitamin A content and bioaccessibility in leaves by combining engineered biosynthesis and storage pathways with high-light treatments. The Plant Journal, 119(6), 2951–2966. https://doi.org/10.1111/tpj.16964
Potrykus, I. (2001). Golden rice and beyond. Plant Physiology, 125(3), 1157–1161. https://doi.org/10.1104/pp.125.3.1157
Rahman, S. U., Khan, M. O., Ullah, R., Ahmad, F., & Raza, G. (2024). Agrobacterium-Mediated Transformation for the Development of Transgenic Crops; Present and Future Prospects. Molecular Biotechnology, 66(8), 1836–1852. https://doi.org/10.1007/s12033-023-00826-8
Sadre, R., Kuo, P., Chen, J., Yang, Y., Banerjee, A., Benning, C., & Hamberger, B. (2019). Cytosolic lipid droplets as engineered organelles for production and accumulation of terpenoid biomaterials in leaves. Nature Communications, 10, 853. https://doi.org/10.1038/s41467-019-08515-4
Sandmann, G., Misawa, N., & Bock, R. (1991). Biosynthesis of carotenoids in microorganisms and plants. European Journal of Biochemistry, 201(1), 1–10.
Shang, A., Gan, R. Y., Xu, X. Y., Mao, Q. Q., Zhang, P. Z., & Li, H. B. (2021). Effects and mechanisms of edible and medicinal plants on obesity: An updated review. Critical Reviews in Food Science and Nutrition, 61(12), 2061–2077. https://doi.org/10.1080/10408398.2020.1769548
Su, Z., Li, J., Zhao, Y., Zhang, T., Zhang, Y., & Liu, J. (2023). Engineering plant lipid droplets for the production of high-value compounds. Molecular Plant, 16(5), 777–794. https://doi.org/10.1016/j.molp.2023.01.008
Tran, Q. G., Cho, K., Kim, U., Yun, J. H., Cho, D. H., Heo, J., Park, S. B., Kim, J. W., Lee, Y. J., Ramanan, R., & Kim, H. S. (2019). Enhancement of β-carotene production by regulating the autophagy-carotenoid biosynthesis seesaw in Chlamydomonas reinhardtii. Bioresource Technology, 292, 121937. https://doi.org/10.1016/j.biortech.2019.121937
Yan, X., Huang, J., Huang, L., Luo, C., Li, Z., Xu, P., Tan, K., Cheong, K.-L., & Tan, K. (2024). Effects of dietary lipids on bioaccessibility and bioavailability of natural carotenoids. LWT, 200, 116171. https://doi.org/10.1016/j.lwt.2024.116171
Zhang, Y., Massel, K., Godwin, I.D., Gao, C. (2018). Applications and potential of genome editing in crop improvement. Genome Biol., 19, 210. https://doi.org/10.1186/s13059-018-1586-y
Zhang, Z., Yu, Z., Liang, D., Song, K., Kong, X., He, M., Liao, X., Huang, Z., Kang, A., Bai, R., & Ren, Y. (2024). Roles of lipid droplets and related proteins in metabolic diseases. Lipids in Health and Disease, 23(1), 218. https://doi.org/10.1186/s12944-024-02212-yZhou, J., Li, D., Wang, G., Wang, F., Kunjal, M., Joldersma, D., & Liu, Z. (2020). Application and future perspective of CRISPR/Cas9 genome editing in fruit crops. Journal of Integrative Plant Biology, 62(3), 269–286. https://doi.org/10.1111/jipb.12793